

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Leptospirosis is a zoonotic disease caused by pathogenic spirochetes in the genus Leptospira [22]. The disease occurs widely in developing countries and is reemerging in the United States [29]. The clinical features are variable and include subclinical infection, a self-limited anicteric febrile illness and severe, potentially fatal disease [22]. In the severe form of leptospirosis (Weil's syndrome), the symptoms include an acute febrile illness associated with multi-organ damage with liver failure (jaundice), renal failure (nephritis), pulmonary hemorrhage, and meningitis [10]. If not treated, the mortality rate may exceed 15% [49]. Furthermore, Leptospira infection can trigger autoimmune diseases in horses as well as humans [36,41]. Several factors associated with virulence have been proposed for Leptospira spp., including the sphingomyelinases, serine proteases, zinc-dependent proteases, collagenase [3], LipL32 [59], the novel factor H-binding protein LfhA [54], and lipopolysaccarides [56].
Pathogenic spirochetes have evolved a variety of strategies to infect host cells such as evasion of the innate as well as adaptive immunity [54]. Attachment to host cells is an essential step for colonization by bacterial pathogens. Leptospira has been shown to bind to mammalian cells, such as Madin-Darby canine kidney (MDCK) cells [2] via the extracellular matrix (ECM) [15]. Several adhesion molecules in the pathogenic spirochetes have been identified including a Fn binding protein (36 kDa protein) [30], a laminin binding protein (Lsa24) [1], and Lig proteins [25,33,34] from Leptospira spp., decorin-binding proteins (Dbp A and B) [37] and Fn-binding proteins (BBK 32 and 47 kDa) [21,38] from Borrelia spp. and MSP, Tp0155, Tp0483, Tp0751 from Treponema spp. [4,5,9]. Lig proteins (Lig A, B and C) possess immunoglobulin-like domains with 90 amino acid repeats that have been identified in other adhesion molecules, such as the intimin of Escherichia coli and the invasin of Yersinia pseudotuberculosis[14,17]. Interestingly, the N-terminal 630 amino acid sequences of LigA and B are identical, but the C-terminal amino acid sequences are variable with only 34% identitify [33]. ligB also encodes a C-terminal, non-repeat domain of 771 amino acid residues [33]. On the other hand, the ligA-ligB intergenic regions from L. kirschneri and L. interrogans are 943 bp and 1347 bp in length respectively, and ligC is not linked to the ligA-ligB locus [25]. The expression of LigA and LigB is controlled by a key environmental signal, osmolarity, to enhance the binding of Leptospira to host cells [26,27].
It has been shown that the lig genes are present exclusively in pathogenic Leptospira spp [25,33]. LigA and LigB are weakly expressed in low passage, but not in high passage cultures of this organism [25,33]. Importantly, we have shown that LigA and LigB expression is upregulated in vivo in the kidneys of Leptospira-infected hamsters [34]. Recently, LigA and LigB have been reported to bind to extracellular matrix proteins including collagens type I and IV, laminin, fibronectin, and fibrinogen [6,24]. These data indicate that Lig proteins may play an important role in attachment of pathogenic leptospires to host cells.
Although there are three copies of lig genes (ligA, B and C) in L. interrogans serovar Pomona and L. interrogans serovar Copenhageni [31,33,34], only ligB is present in most pathogenic Leptospira spp. ligA is absent in L. interrogans serovar Lai [42], ligC is truncated (a pseudogene) in L. kirschneri serovar Grippotyphosa [25] and both ligA and ligC are absent in L. borgpetersenii serovar Harjo [3]. Therefore, we focused on LigB in this study and report that the variable region of LigB binds with high affinity to Fn, suggesting that this fragment is crucial for bacterial adhesion to host cells.
Materials and Methods
Bacterial strains and cell culture
L. interrogans serovar Pomona (NVSL1427-35-093002) was used in this study [35]. All experiments were performed with virulent, low-passage strains obtained by infecting golden syrian hamsters as previously described [35]. Leptospires were grown in EMJH medium at 30℃ for less than 5 passages and growth was monitored by darkfield microscopy. The MDCK cells (ATCC CCL34) were cultured in Dulbecco minimum essential medium containing 10% fetal bovine serum (GIBCO, USA) and were grown at 37℃ in a humidified atmosphere with 5% CO2.
Reagents and antibodies
Horseradish peroxidase (HRP)-conjugated goat anti-hamster antibody, HRP-conjugated goat anti-mouse antibody and HRP-conjugated goat anti-rabbit antibody were purchased from Zymed (USA). Rabbit anti-glutathione S-transferase (GST) antibody, Alexa 594-conjugated goat anti-hamster antibody, Alexa 488-conjugated goat anti-hamster antibody, and FITC-conjugated goat anti-mouse antibody were purchased from Molecular Probe (USA). Anti-Fn (MAB1932) and anti-actin mouse antibodies (MAB1501) were purchased from Chemicon International (USA). Human plasma Fn was purchased from GIBCO (USA). Anti-L. interrogans antibodies were prepared in hamsters as previously described [35].
Plasmid construction and protein purification
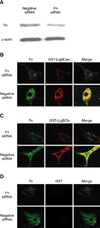
Constructs for the expression of GST, GST fused with the conserved region of LigB (LigBCon; amino acids 1-630) and GST fused with the central variable region of LigB (LigBCen; amino acids 631-1417) were previously generated using the vector pGEX-4T-2 (Amersham Pharmacia Biotech, USA) [33]. GST fused with the C-terminal variable region of LigB (LigBCtv; amino acids 1418-1889) was generated using the vector pET41A (Novogen, USA). Relevant fragments of DNA were amplified by PCR using primers based on the ligB sequence [33]. Primers were designed to introduce a SalI site at the 5' end of each fragment and a stop codon followed by a NotI site at the 3' end of each fragment. The PCR products were digested sequentially with SalI and NotI and then ligated into pGEX-4T-2 or pET41A cut with SalI and NotI. We purified the soluble form of GST-LigBCon, GST-LigBCen and GST-LigBCtv from E. coli as previously described [34,35].
Binding assays by ELISA
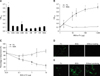
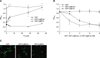
To measure the binding of Leptospira to the ECM components, 1 mg of each ECM component (as indicated in Fig. 1A) in 100 µl PBS (pH 7.2) was coated onto microtiter plate wells. For the dose-dependent binding experiments, different concentrations of Fn (as indicated in Fig. 1B) were coated onto the microtiter plate wells. The plates were incubated at 4℃ for 16 h and subsequently blocked with blocking buffer (50 µl/well) containing 3.5% BSA in 50 mM Tris (pH 7.5)-100 mM NaCl-1 mM MgCl2, MnCl2, and CaCl2 at room temperature (RT) for 2 h. Then, the Leptospira (107) were added to each well and further incubated at 37℃ for 6 h. To determine the inhibition of Leptospira binding to the MDCK cells by Fn, the Leptospira (107) were pre-incubated at 37℃ for 1 h with various concentrations of Fn (as indicated in Fig. 1C) prior to the addition of the MDCK cells (105) and finally incubated for 6 h at 37℃. The percentage of adhesions was determined relative to the attachment of the untreated Leptospira binding to the MDCK cells. For all experiments, the same concentration of BSA was used as a negative control. To determine the binding of LigBCen or LigBCtv to Fn, 10 nM of GST-LigBCen, GST-LigBCtv or GST (negative control) was added to 96 well microtiter plates coated with various concentrations of Fn (as indicated in Fig. 3A) or BSA (negative control and data not shown) in 100 µl PBS for 1 h at 37℃.
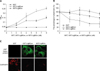
To measure the binding inhibition of Leptospira to Fn, various concentrations of GST-LigBCen, GST-LigBCtv (as indicated in Fig. 3B) or GST (negative control) in 100 µl PBS was added to Fn or BSA (negative control and data not shown) (1 mg in 100 µl PBS) coated wells at 37℃ for 1 h, then the Leptospira (107) were added to each well and incubated at 37℃ for 6 h. To measure the binding of LigBCen or LigBCtv to the MDCK cells, the MDCK cells (105) were incubated with various concentrations (as indicated in Fig. 4A) of GST-LigBCen, GST-LigBCtv or GST (negative control) in 100 µl PBS for 1 h at 37℃. To measure the binding inhibition of Leptospira to the MDCK cells treated with LigBCen or LigBCtv, the MDCK cells (105) were pretreated with various concentrations (as indicated in Fig. 4B) of GST-LigBCen, GST-LigBCtv or GST (negative control) in 100 µl PBS for 1 h at 37℃. Then, the Leptospira (107) were added to each well and incubated for 6 h at 37℃. Following the incubation, the plates were washed three times with phosphate-buffered saline (PBS) containing 0.05% Tween-20 (PBST). To measure the binding of the Leptospira, hamster anti-Leptospira (1:200) and HRP-conjugated goat anti-hamster IgG (1:1,000) were used as primary and secondary antibodies, respectively. To detect the binding of GST-LigBCen, GST-LigBCtv, or GST to Fn or the MDCK cells, rabbit anti-GST (1:200) and HRP-conjugated goat anti-rabbit IgG (1:1,000) were used as primary and secondary antibodies, respectively. After washing the plates three times with PBST, 100 µl of TMB (KPL, USA) was added to each well and incubated for 5 min. The reaction was stopped by adding 100 µl of 0.5% hydrofluoric acid in each well. Each plate was read at 630 nm by an ELISA plate reader (Bioteck EL-312; BioTeck, USA). Each value represents the mean ± standard error of the mean (SEM) of three trials performed in triplicate samples. Statistically significant (p < 0.05) differences are indicated by asterisks.
Binding assays by epifluorescence microscopy (EPM) and confocal laser-scanning microscopy (CLSM)
To measure the binding of Leptospira to Fn by EPM, Leptospira (108) were added to each well (eight well culture slides) coated with 1 mg Fn or BSA (negative control) in 100 µl of PBS and incubated at 37℃ for 6 h (Fig. 1D). To measure the binding inhibition of Leptospira to the MDCK cells by Fn, 108 Leptospira were pre-incubated with 10 µg of Fn or BSA (negative control) in 100 µl of PBS for 1 h at 37℃ prior to the addition of 106 MDCK cells and incubated 6 h at 37℃ (Fig. 1E). To measure the binding inhibition Leptospira to Fn by LigBCen or LigBCtv by EPM, 50 nM of GST-LigBCen, GST-LigBCtv or GST (negative control) in 100 µl PBS was added to each of the Fn or BSA (negative control and data not shown) (1 mg per 100 µl) coated wells for 1 h at 37℃. Then, the Leptospira(108) were added to each well and incubated for 6 h at 37℃ (Fig. 3C). To determine the binding inhibition of Leptospira to the MDCK cells by LigBCen or LigBCtv by CLSM, the MDCK cells (106) were preincubated with 50 nM of GST-LigBCen, GST-LigBCtv or GST (negative control) in 100 µl of PBS for 1 h at 37℃ respectively. Then, the Leptospira (108) were added to each well and incubated for 6 h at 37℃ (Fig. 4C). For the detection of Leptospira binding in Figs. 1D, E, and Fig. 3C, hamster anti-Leptospira antibodies (1:100) and Alexa 488-conjugated goat anti-hamster IgG (1:250) were used as primary and secondary antibodies, respectively. To determine the attachment of Leptospira and the binding of GST-LigBCen, GST-LigBCtv or GST, Fig. 4C, rabbit anti-GST (1:250) and hamster anti-Leptospira antibodies (1:100) served as primary antibodies, and FITC conjugated goat anti-rabbit IgG (1:250) and Alexa 594-conjugated goat anti-hamster IgG (1:250) were used as secondary antibodies. Fixation and immunofluorescence staining were performed as previously described [44] with slight modifications. Briefly, Leptopsira and the MDCK cells were fixed in 2% paraformaldehyde for 60 min at RT. For the antibody labeling, fixed bacteria were incubated in PBS containing 0.3% BSA for 10 min at RT. The primary and secondary antibodies, in the PBS containing 0.3% BSA, were incubated sequentially for 60 min at RT. After incubation with the primary and secondary antibodies, the glass slides were mounted with coverslips using Prolong Antifade (Molecular Probe, USA) and viewed with a 60 × objective by EPM (Nikon, Japan) or CLSM (Olympus, Japan). An Olympus Fluoview 500 confocal laser-scanning imaging system, equipped with krypton, argon and He-Ne lasers on an Olympus IX70 inverted microscope with a PLAPO 60 × objective, was used. The settings were identical for all captured images. Images were processed using Adobe Photoshop CS2. For counting the attachment of Leptospira to the MDCK cells or Fn, three fields were selected to count the number of binding organisms. All studies were repeated three times and the number of Leptospira attached to the MDCK cells were counted by an investigator blinded to the treatment group.
GST pulldown assay
The GST pull-down assay was performed as previously described [57]. Purified proteins or GST (negative control) were loaded onto 0.5 ml glutathione-Sepharose beads (Amersham Biosciences Piscataway, USA) at 4℃ overnight. The beads were then washed three times with the lysis buffer containing 30 mM Tris acetate, 10 mM sodium phosphate, pH 7.4, 0.1% Tween 20, 1 mM EDTA, 2 µg/ml leupeptin, 4 µg/ml aprotinin, 1 µg/ml pepstatin A, and 1 mM phenylmethylsulfonyl fluoride (PMSF). The MDCK cells (106) were lysed in the lysis buffer and used immediately after lysis. A 500 µl aliquot of cell lysate or human plasma Fn (40 µg/ml) was incubated with purified proteins immobilized on glutathione-Sepharose beads at 4℃ for 3 h. After incubation, the beads were separated by centrifugation, washed three times with the lysis buffer and boiled in Laemmli sample loading buffer consisting of 50 mM Tris-HCl (pH 6.8), 100 mM dithiothreitol, 2% sodium dodecyl sulfate, 0.25 mM PMSF, and 0.1% bromophenol blue in 20% glycerol. The eluted proteins were subjected to 6% SDS-PAGE and electroblotted onto polyvinylidene difluoride membranes. The membranes were incubated in 5% skim milk in PBS/T overnight and then incubated with mouse anti-Fn antibody (1:1,000). The immunocomplexes were detected with an HRP-conjugated goat anti-mouse IgG antibody (1:5,000).
Small interfering RNA (siRNA) inhibition of LigB binding
The siRNA duplexes directed against the sequence 5'-gcagcacaacuuccaauua-3' of Fn and negative siRNA duplex, 5'-auucuaucacuagcgugac-3', were selected by the software, siDESIGN [43] and synthesized by Dharmacon (USA). The RNA duplexes were introduced into the MDCK cells by the method of lipofection [18], and 8 × 105 cells were transfected with 0.4 µg negative siRNA and Fn-siRNA. Adhesion assays were performed 72 h after lipofection [51]. The knockdown efficiency of endogeneous Fn expression was determined as previously described [57] with slight modification. The total protein contents of the MDCK cells (106) were analyzed using Western immunoblotting as described under 'GST pulldown assays'. The protein bands of actin derived from the MDCK cells were measured as a control using a mouse anti-actin antibody (1:5,000). The band intensity was measured by densitometry using the Image J software (National Institutes of Health, Bethesda, MD, USA) [53]. A LigB binding assay was performed 72 h after lipofection. To determine the binding of LigB fragments to Fn, each fragment (50 nM) was added to the MDCK cells (106) transfected with Fn or negative siRNA. To determine the binding of each fragment and the expression of Fn in the MDCK cells, rabbit anti-GST (1:250) and mouse anti-Fn (1:250) served as the primary antibodies, and FITC-conjugated goat anti-mouse IgG (1:250) and Texas Red-conjugated goat anti-rabbit IgG (1:250) were used as secondary antibodies. Fixation, immunofluorescence staining, image detection, and processing were carried out as described in previous sections. All experiments were performed in triplicate.
Isothermal titration calorimetry
The experiments were carried out with CSC 5300 microcalorimeter (Calorimetry Science, USA) at 25℃ as previously described [47]. In a typical experiment, the cell contained 1 ml of a solution of proteins, and the syringe contained 250 µl of a solution of Fn at a concentration that was 20 times higher than the protein concentration in the cell. Both solutions were in PBS pH 7.5. The titration was performed as follows: 15 to 25 injections of 10 µl (Table 1) with a stirring speed of 250 rpm, and the delay time between the injections was 5 min. Data were analyzed using Titration BindingWork 3.1 software (Calorimetry Science, USA) that was fit to an independent binding model. The concentration of Fn and LigB used in this study was based on our preliminary titration experiments (data not shown).
Statistical analysis
Statistically significant differences between samples were determined using the Student's t-test following logarithmic transformation of the data. Two-tailed p-values were determined for each sample, and a p < 0.05 was considered significant. Each data point represents the mean ± SE of a sample tested in triplicate. An asterisk indicates that the result was statistically significant.
Results
Attachment of Leptospira to the MDCK cells was mediated by fibronectin
The binding of leptospiral cells to various ECM components was determined by ELISA. As shown in Fig. 1A, Leptospira were strongly bound to Fn, but not to other ECM molecules (Fig. 1A). Furthermore, the binding of Leptospira to Fn was dose dependent (Fig. 1B). When Leptospira were pretreated with Fn, binding to the MDCK cells was decreased (Fig. 1C). There was an approximately 3.5-fold increase in the immobilization of Leptospira in the Fn-coated wells compared to the controls (Fig. 1D). Moreover, Fn was observed to block the attachment of Leptospira, by approximately 47%, when the Fn treated Leptospira were added to the MDCK cells (Fig. 1E). Thus, Fn appears to mediate the attachment of Leptospira to the MDCK cells.
Interaction between LigB and Fn
To determine whether LigB interacts with Fn, we truncated the LigB protein into three parts, LigBCon, LigBCen and LigBCtv, (Fig. 2A) due to the difficulty of expressing and purifying the full length LigB [33]. First, we analyzed the interaction of each LigB fragment with Fn using a GST-pull down assay. Our results showed that both human plasma Fn and Fn derived from the MDCK cell lysates could bind both LigBCen and LigBCtv, but not LigBCon (Figs. 2B and C). Since LigBCen and LigBCtv showed a positive pull down result, the interaction between LigBCen and LigBCtv with Fn was further studied by ELISA. We found that both the binding of LigBCen and LigBCtv to Fn, and the inhibition of the attachment of Leptospira to Fn by LigBCen and LigBCtv, were dose-dependent (Figs. 3A and B). Moreover, the EPM images revealed an up to 40% reduction in the attachment of Leptospira to Fn in the presence of LigBCen and LigBCtv (Fig. 3C). Finally, in order to quantitatively evaluate the binding affinity between Fn and LigB fragments, the dissociation constants (Kd) were measured by ITC (Table 1). Fig. 4 shows the data from a typical ITC experiment. The interaction appears to be exothermic with a favorable enthalpy and unfavorable entropy. The calculated Kd values for Fn binding to LigBCen and LigBCtv were 0.01 µM and 8.55 µM, respectively (Table 1). The binding of LigBCon could not be detected by ITC (data not shown). These findings are in agreement with our previous results. Altogether, these data indicate that Fn specifically interacts with LigBCen and LigBCtv fragments.
LigBCen and LigBCtv mediate the attachment of Leptospira to the MDCK cells
To determine if LigB is used by Leptospira to adhere to the MDCK cells, various concentrations of LigBCen or LigBCtv were added to the MDCK cells, and binding was detected by ELISA and immunofluorescence staining. Our results clearly showed that LigBCen and LigBCtv were bound to the MDCK cells in a dose dependent manner (Fig. 5A). Pretreatment of the MDCK cells with LigBCen or LigBCtv reduced the attachment of Leptospira by ~32%. The reduction of Leptospira attachment was also dose-dependent (Figs. 5B and C). We further elucidated the receptor role of Fn in the MDCK cells for its possible ligand, LigB on the surface of Leptospira, by RNA interference to decrease the endogeneous Fn expression in the MDCK cells. As shown in Fig. 6A, transfection of the cells with siRNA duplex specific for canine Fn resulted in a ~36% reduction of the Fn expression, relative to the control cells. The binding of LigBCen and LigBCtv to Fn siRNA-transfected MDCK cells was significantly reduced (Figs. 6B and C). These results suggest that Fn serves as a receptor for LigB that mediates Leptospira adhesion.
Discussion
Adhesion to host cells is pivotal for many pathogenic bacteria including Leptospira spp. Since pathogenic Leptospira spp. can infect a variety of tissues including liver, kidney and lung, study of the host-pathogen interaction is extremely important for improved understanding of leptospirosis. Recently, the leptospiral genome has been sequenced and a number of tentative virulence factors have been proposed [3,31,42]. However, their exact roles in leptospiral pathogenesis remain to be established. To date, several leptospiral adhesion molecules have been identified. These include a 36 kDa Fn-binding protein [30], a 24 kDa laminin-binding protein [1] and LigA, LigB and LigC proteins [25,33,34]. These molecules may play an important role in the pathogenesis of leptospiral infection since they are able to bind to ECMs such as collagens I and IV, laminin and fibronectin [6,24].
Pathogenic Leptospira spp. have been previously reported to adhere to extracellular matrices [15,16] including Fn. Fns are dimers of two similar peptides linked at their C-termini by two disulfide bonds [8] and serve as receptors for several bacteria, including spirochetes [7,11,12,19,20,23,28,32,38,40,46,50,55]. Our results showed that Fn immobilized Leptospira. In addition, Fn was observed to block the attachment of Leptospira to MDCK cells if the Leptospira were pre-treated with Fn. These results support the recent report that Fn might be an important molecule involved in the pathogenic adherence of Leptospira spp. to host cells [6,24].
We demonstrated the interaction between LigB and Fn. It was shown that the LigBCen and LigBCtv fragments were bound to Fn, by GST-pulldown assays, ELISA and ITC measurements. The low Kd values for LigBCen indicated that the LigB-Fn interaction was specific. This evidence strongly suggests that LigB is a Fn-binding protein. A study reported by Choy et al. [6] showed that LigB U1 and LigB U2 (LigBCen equivalent) could strongly bind to Fn, while the LigB CTD (LigBCtv equivalent) binds weakly to Fn. However, the Kd values of LigBCen and LigBCtv to Fn that we obtained were slightly different than those reported by Choy et al. [6]. The differences in the obtained Kd values could be explained by (i) the protein fragments evaluated in this study (LigBCen and LigBCtv) were not exactly the same length fragments (LigBU1, LigBU2 and LigBCTD) and (ii) the method we used (ITC) to measure the Kd differed from that of Choy et al. [6].
Since pathogenic Leptospira spp. adheres to renal tubular epithelial cells and induces a severe tubulointerstitial nephritis leading to renal failure [58], it is possible that LigB is responsible for the binding of Leptospira to the renal tubular epithelium. Our results indicated that LigB binds to the MDCK cells via the LigBCen or the LigBCtv fragments. However, the LigBCen was observed to bind to both the MDCK cells and Fn with a greater affinity than the LigBCtv. The microscopic images also showed that not all of the Fn was co-localized with the LigB. This result suggests that LigB might bind to two or more receptors. Our results elucidate the process of Leptospira attachment to the MDCK cells, as noted in a previous study [52], and demonstrated how Fn can block leptospiral attachment to the MDCK cells.
Our results clearly confirm that LigB is one of the microbial surface components that recognize adhesive matrix molecules (MSCRAMM) members that bind to the ECM including Fn. The transmembrane domain of LigB is predicted to reside within the conserved region, with only the variable region exposed on the surface [33,34]. These results support our data that Fn-binding domains of LigB are localized in the variable regions. This is not surprising since similar findings have been reported for other MSCRAMMs [13,37,39]. In Borrelia, the binding motifs in the decorin-binding proteins, DbpA and B, are located in the central regions, which vary among the different Borrelia strains (B. burgdorferi, B. garnii, and B. afzeli) [37]. The Fn-binding domain of the Fn-binding protein, BBK32 is also variable among the different Borrelia strains [39]. The repetitive D1, D2 and D3 elements of Staphylococcus aureus Fn-binding protein, which bind the N-terminal 29 kDa of Fn, also vary [13].
Since both LigBCen and LigBCtv bind to Fn, but with different affinities, this suggests that there is more than one potential Fn-binding domain. In Mycobacterium avium, two Fn-binding domains are located on two non-contiguous segments of 24 amino acids in the Fn attachment protein-A [45]. The FnBPA of Staphylococcus aureus contains three repetitive elements, D1, D2 and D3 and each binds the N-terminal 29 kDa fragment of Fn [13]. Seven additional Fn-binding elements are located in the N-terminal of the D repeats [48]. In Streptococcus dysgalactiae, there are five Fn-binding segments within the C-terminus of the Fn binding protein F1/(FnBB) [47,48]. Therefore, it is likely that several binding sites might be present in the LigB variable region. However, we were unable to identify a similar Fn-binding motif in the other known Fn-binding proteins.
In conclusion, we have shown that LigBCen and LigBCtv bind to Fn and have confirmed that LigB is a member of the MSCRAMMs. Since pathogenic Leptospira spp. initially attaches to mucosal epithelial cells prior to entry into the bloodstream and subsequent dissemination to multiple organs such as the kidney, liver and lung, Lig proteins may play a pivotal role in the pathogenesis of leptospirosis. Fn is one of the most important ECMs on epithelial cells and serves as a receptor for leptospiral adherence [6,15,24]. Thus, further studies into the interaction of Lig proteins and ECMs are warranted.



 XML Download
XML Download