

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Parkinson's disease (PD) is characterized by a loss of dopamine in the striatum resulting from the progressive degeneration of dopaminergic neurons within the substantia nigra [20]. Tyrosine hydroxylase (TH) is the rate-limiting enzyme in the synthesis of catecholamine, and is widely used as a marker of dopaminergic cells [23]. Tang et al. [32] have shown that dopaminergic differentiation of striatum-derived precursor cells following transplantation requires TH induction. Furthermore, Lu et al. [16] observed significant functional recovery in rats model of PD receiving TH-engineered mesenchymal stem cells transplanted into the striatum. Moreover, Peschanski et al. [25] have demonstrated bilateral motor improvement in two patients with PD after grafting of fetal mesencephalic tissue. Therefore, enhancing TH gene expression or protein production is a critical step for treatment of PD.
The PC12 cell line is a neural crest-derived adrenal chromaffin cell line obtained from a rat pheochromocytoma. PC12 cells serve as a useful model system for studying the molecular mechanisms responsible for sympathetic neuronal differentiation induced by nerve growth factor (NGF) because they can synthesize and store the catecholamine neurotransmitters dopamine and norepinephrine [8]. Mesenchymal stem cells (MSCs) derived from bone marrow can differentiate into osteoblasts, chondrocytes, adipocytes [26], and neural cells [15,24]. In addition, MSCs express neuroregulatory molecules, such as NGF, brain-derived neurotrophic factors (BDNF), and glial cell-derived neurotrophic factors (GDNF) [5,6,14,17,18]. NGF can upregulate TH gene expression in PC12 cells [7,13]. Others have reported that induction of TH gene expression can be achieved by co-treatment fetal cerebral cortex with dopamine and BDNF [37,38]. Therefore, novel methods of inducing TH expression via treatment with growth factors, either directly or through cell co-cultures, has significant potential.
Co-cultures of two different cell types provide insight into mechanisms of cell-cell interactions. Direct-contact co-cultures can promote more mature differentiation of neural stem cells [30], which also involves the extracellular matrix or soluble factors secreted by supporting cells [1,30,31]. Moreover, direct contact cultures better mimic intracerebral grafting in terms of the cell-cell interactions.
The cat is the preferred model for studying human neurodegerative disorders [3,19,27,36]. It provides a number of important advantages for biomedical research, particularly stem cell transplantation in fetal and postnatal animals. The purpose of this study was to explore whether MSCs derived from feline bone marrow could enhance the expression of TH in PC12 cells by direct contact co-cultures.
Materials and Methods
Isolation and culture of feline MSCs
The protocol used in this study was approved by the Gyeongsang National University Animal Use Committee. Feline bone marrow was harvested by bone marrow aspiration from a greater trochanter of the femur. Mononuclear cells were isolated from the collected samples by the Ficoll 400 (Sigma, USA) density gradient method. The cells were then rinsed twice with PBS, and grown in Dulbeco's Modified Eagle high glucose medium (DMEM, 4.5 g/l glucose; Gibco-BRL, USA) with 10% fetal bovine serum (FBS; Gibco-BRL, USA) and penicillin/streptomycin (50 IU/ml / 50 µg/ml; Sigma, USA). After 5 days, the resulting monolayer of cells, hereafter named bone marrow-derived MSCs, was trypsinized and aliquots were frozen and stored or cultivated further.
Co-culture of PC12 cells and MSCs
MSCs were from passage 4. The study was divided into three experimental groups: a MSC group, a PC12 cell group, and the combined MSC + PC12 cell group. The cells were seeded at 1.5 × 105 cells of each cell type (3 × 105 total in the combined group) onto 60 mm dishes and grown in DMEM supplemented with 10% FBS and penicillin/streptomycin (50 IU/ml / 50 µg/ml). Cells were incubated at 37℃ in a humidified atmosphere of 5% CO2 for 3 days.
Flow cytometric determination of cell-surface antigen profile
Ten microliters of CD9 (Serotec, USA), CD18, CD44, and CD45-like (VMRD, USA) cat-specific antibodies were added to the bottom of tubes, followed by 100 µl of 6 × 106 cells/ml single cell suspensions of cultured MSCs in the 4th passage. The mixture was incubated for 30 min at 4℃ in the dark and washed. Five microliters fluorescent-tagged secondary antibodies were then added to the resuspended cells for 30 min at 4℃ in the dark. Positive cells were then detected on flow cytometers.
Immunocytochemistry of MSCs and TH expression in PC12 cells
To identify MSCs from feline derived bone marrow, the primary chicken anti-vimentin antibody (1 : 5,000, polyclonal; Chemicon, USA) was employed; the secondary antibody was a FITC-conjugated antibody against chicken IgG (1 : 320; Sigma, USA). To identify TH-positive differentiation of feline MSCs in co-cultures experiment, MSCs were prelabeled using Dil-Ac-LDL (1 : 20; Biomedical Technologies, USA) and then co-cultured with PC12 for three days. Anti-TH (1 : 500, polyclonal; Chemicon, USA) was the primary antibody used to identify TH in all groups; a FITC-labeled antibody against rabbit IgG (1 : 80; Sigma, USA) was used as the secondary antibody. The cells were cultured in a four-well chamber slide and fixed with 4% paraformaldehyde for 10 min. Following fixation, the cells were treated with 0.3% Triton X-100 containing 10% normal goat serum for 30 min. Cells were then incubated with the primary antibody at room temperature for 2 h, followed by incubation with the FITC-conjugated secondary antibody at 37℃ for 30 min. To prevent non-specific staining, secondary antibodies were used at the same concentrations as the primary antibodies. The slides were examined with an microscope (Olympus, Japan) equipped with fluorescent illumination.
Western blots of TH protein
Western blot analysis was performed to determine whether the presence of MSCs affected TH protein expression in PC12 cells. The cells were lysed using a SDS-sample buffer. After ultracentrifugation (12,000 rpm/4℃/10 min), the supernatant was collected. Fifteen µg of protein was separated by SDS-PAGE and transferred to a PVDF membrane (Bio-Rad, USA). Immunoreactions were performed with antibodies against TH (1 : 500, polyclonal; Chemicon, USA) and GAPDH (1 : 300, monoclonal; Chemicon, USA). The membrane was washed and incubated with horseradish peroxidase conjugated anti-rabbit IgG (1 : 1,000; Santa Cruz Biotechnology, USA) or anti-mouse IgG (1 : 5,000; Chemicon, USA). Antigen protein was visualized using chemiluminescence with an ECL-detecting reagent (Amersham Pharmacia Biotech, USA) according to the protocol of the manufacturer.
Statistical analysis
Experiments were repeated three times. The band from the western blot was scanned and analyzed by densitometry using the Sigma Gel System (SPSS, USA). Density values are expressed as the mean ± SD. Statistical analysis was performed using one-way ANOVA analysis followed by a posthoc LSD test. The acceptance level for statistical significance was p<0.05.
Results
Establishment of primary culture
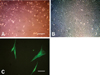
The feline bone marrow-derived mononuclear fraction was isolated and then cultured. Attached cells were observed at 10-14 days after the initial plating. These cells comprised a morphologically heterogeneous population of cells that included spindle-shaped cells, large flat cells, and small round cells (Fig. 1A).
Cell-surface antigen profile
The feline MSC-surface antigen profile was ascertained by staining with feline-specific monoclonal antibodies followed by flow cytometry (Fig. 2). Feline MSCs were strongly positive for CD9 and CD44, but negative for CD18 and CD45. They demonstrated typical characteristics of mesenchymal cells, including the capacity to transform into osteocytes and adipocytes.
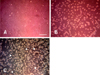
Morphology in co-culture of PC12 cells and MSCs
In the MSCs alone, the cells showed an elongated fibroblast shape (Fig. 3A). In the PC12 cells alone, the cells showed modest growth and tended to grow in small clusters (Fig. 3B). In the co-cultured group, both cells appeared robust. Although the PC12 cells did not change morphology significantly, the MSCs possessed long, thin, complex processes (Fig. 3C).
Western blot analysis of TH expression in PC12 and MSC co-cultures
Representative immunoblot bands for TH proteins are shown in Fig. 4. TH protein demonstrated a stronger signal at 60 kDa in the co-culture group than the PC12 cell group (p<0.001). In contrast, TH wasn't expressed in the MSC alone group.
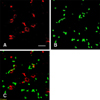
Immunocytochemistry of TH expression in PC12 and MSC co-cultures
MSCs (red) did not show immunostaining of TH protein (green) (Figs. 5A-C), indicating that the MSCs had not differentiated into TH positive cells under our experimental conditions.
Discussion
Here, we show that MSCs derived from feline bone marrow can enhance TH protein expression in PC12 cells. There are several possibilities for what mediates this induction. First, PC12 cells can synthesize and release dopamine [8], and MSCs can secrete NGF and BDNF [5]. NGF can up-regulate TH mRNA expression in PC12 cells [13], and TH gene expression can be induced in human and murine fetal cerebral cortex by co-treatment with dopamine and BDNF [37,38]. Therefore, the NGF and BDNF produced by MSCs and the dopamine from PC12 cells could stimulate TH induction [13,37,38].
Second, PC12 cells originated from adrenal chromaffin cell-derived tumors [8]. Adrenal medullary chromaffin cells produce FGF-2, TGF-β, GDNF, etc [11,35]. Adult rat bone marrow stromal cells express genes which are associated with dopaminergic neurons [10], and MSCs can differentiate into dopaminergic neurons both in vitro and in vivo [12,33,34]. Here, MSCs did not express TH in western blots or immunostaining, perhaps due to immaturity of the cells at the time of the immunocytochemical study.
Third, studies in both chromaffin and PC12 cells reveal that TH gene expression is dependent on cell density [2,8,28,29]. The increase in TH mRNA at a high cell density is correlated with higher levels of TH immunoreactive protein, increased enzymatic activity, and elevated dopamine content [4,8]. However, TH expression did not vary with increasing PC12 cell density (data not shown), possibly due to the short-term culture conditions. Chromaffin cell differentiation was also not affected by increased cell density [30].
Fourth, direct cell-cell contact may help enhance TH expression. Direct contact between bone marrow stromal cells and the host brain tissue is important for differentiation [1]. Direct chromaffin cell contact may stimulate differentiation of neuronal progenitor cells [30]. The direct interaction of MSCs with neurons promoted long-last survival [31].
In summary, co-culture of MSCs with PC12 cells increases TH expression in the cultures. In vivo, MSCs from bone marrow markedly increase the proliferation of hippocampal-derived neural stem cells via secretion of cytokines [22]. Furthermore, MSCs transplantation into rats following traumatic brain injury improved functional recovery [15]. However, only a small number of transplanted cells expressed neuronal antigens in that study, making functional improvement in recipients unlikely. Therefore, the secretion of neurotrophic factors or direct cell-cell contact by MSCs may be more important than the resulting neural differentiation in transplant experiments. Our results imply a new role for MSCs in treating neurodegenerative diseases such as PD.

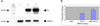
 XML Download
XML Download