

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Sertoli cells play an important role in determining the testis size, germ cell number and sperm production rate in adulthood [2]. The number of spermatozoa produced per day is also critically dependent on the number of Sertoli cells per testis because each Sertoli cell can support only a finite number of germ cells [13]. In all species studied so far, Sertoli cells proliferate during fetal or neonatal life as well as in the peripubertal period [16]. The period for the proliferation of Sertoli cells has been well established in certain mammals. In rats, it has been found to commence during the neonatal period and cease at 18 days of age [12]. In bulls, boars and rams, similar time spans have been observed but Sertoli cell replication has been found to last from 6 to 10 weeks [3,4,9,15]. Germ cells proliferate after a short period of quiescence and migrate from their central location towards the basal membrane, at which stage they are known as spermatogonia [10,11]. Knowledge of when Sertoli cell proliferation ceases in a species is important for planning studies and correctly interpreting the results. However, there has been no direct determination of these periods in birds, even the domestic fowl. The aims of this study were observe the cessation of post-hatching mitotic proliferation of Sertoli cells in domestic fowl, and to determine the volume density of Sertoli and germ cells during this period.
Materials and Methods
White Leghorn (Lohmann LSL) male chicks were identified by a cloacal inspection and kept under a 14Light : 10Dark lighting regime. The chicks were provided with food and water ad libitum. Five animals were selected randomly every week between one and 10 weeks of age. They were injected intraperitoneally with 5-bromo-2-deoxyuridine (BrdU, 100 mg/kg body weight), as a specific marker for cellular proliferation, and 1 h later with heparin (130 IU/kg). The chicks were then anesthetized with CO2 and perfused through the heart with a Bouin's solution using a peristaltic pump (Ismatec, Switzerland) for 10-20 min [18]. A complete color change of the liver to yellow was considered the criterion for ending perfusion. This method, cardiac perfusion with heparin, resulted in one testis not being well fixed. Other studies have used this method for rat testis perfusion with an average 99% success rate [14]. Moreover, it is easily performed and does not require experienced users.
The left testis was taken from the animals, immersed in the same fixative for 24 h and embedded in paraffin. Four-micron thick specimens were taken from the testes in a random systematic manner.
The sections were immunostained for BrdU to display the cellular proliferation. The sections were deparaffinized, rehydrated and treated with 0.3% H2O2 for 10 min in methanol. Antigen retrieval was achieved by incubating the slides with trypsin (Lab Vision, USA) for 20 min. The sections were washed with PBS (3 × 5 min) and incubated with 2 N HCl for 30 min and then with a 0.1 M borax solution for 10min. Following washing with PBS (3 × 5min), the sections were incubated for 5 min with Ultra V Block (Lab Vision, USA) to block the non-specific binding, and incubated with the anti-BrdU primary antibody (Lab Vision, USA) for 2 h at room temperature. The immunoreaction was detected using a detection kit (UltraVision; Lab Vision, USA). This involved incubating the sections with biotinylated anti-mouse immunogloblins and streptavidin peroxidase for 10 min each, and visualizing any immunoreactivity using a 3,3'-diaminobenzidine as chromagen substrate. The sections were then counterstained lightly with Mayer's hematoxylin.
In order to determine the volume density using the standard point counting method [1], testes sections from 5 animals each week were stained with Harris' hematoxylin and eosin, and observed by projecting objective images (×100) onto a monitor. Ten fields were evaluated using a systematic clock-face sampling pattern from a random starting point. This involved placing a 104-point grid printed on a transparency over each field and counting the number of grid points over the Sertoli cell nuclei, germ cell nuclei or seminiferous tubule. The total number of points over each structure, adding all 10 fields, was obtained. The total number (N) of grid points over the seminiferous tubules is expressed as the percentage of the total number of grid points (= 1040). For the Sertoli and the germ cell nuclei, the total number of grid points over each structure was divided by N and is expressed as a percentage.
An independent samples t-test was used to evaluate the differences between the mean volume density of Sertoli cell and germ cell nuclei within the same week. A one-way ANOVA test was used to examine the changes in the mean volume densities of Sertoli cells, germ cells and seminiferous tubules with time.
Results
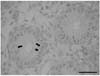
Fig. 1 shows the clear differentiation of Sertoli cells and germ cells in a typical immature testis. The germ cells are spherical with large nuclei. The Sertoli cell nuclei are considerably smaller with an irregular shape.
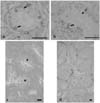
BrdU labeling was strong and without a background, making it particularly suitable for studies of domestic fowl testes. Clear labelling of the Sertoli and germ cells was observed between one and seven weeks of age. However, at 8 weeks, Sertoli cell labeling was not observed in 3 fowls (Fig. 2a) but was observed in the remaining 2 fowls (Fig. 2b).
An examination of the hematoxylin stained sections at 8 weeks revealed primary spermatocytes to be present in all testes in which the Sertoli cells had not been labeled (Fig. 2d). Indeed, no primary spermatocytes were observed in any sections in which the Sertoli cells were labelled (Fig. 2c). Moreover, spermatagonia were the only cells labelled for BrdU after week 8. This suggests that the cessation of Sertoli cell replication coincides with the commencement of meiosis.
The volume density of the seminiferous tubules increased until week 8 with a minor decrease observed at weeks 2 and 5 (Fig. 3). The volume density of the germ cell nuclei was initially less than that of the Sertoli cell but these volume densities converged becoming virtually the same by week 6, and remaining relatively constant until the commencement of spermatogenesis (Fig. 4).
Discussion
Although there is no knowledge of the precise period and kinetics of Sertoli cell proliferation in domestic fowl, hemicastration studies have provided some information. Compensatory testicular hypertrophy of the testis after hemicastration has been observed to occur for up to 8 weeks of age [5,7]. Because the testis size correlates with the number of Sertoli cells number adulthood, it has simply been assumed that the Sertoli cell proliferation period occurs during the first 8 weeks post-hatching. The results from this study confirm this assumption. Therefore, it is recommended that any research involving the testes of domestic fowl should consider that Sertoli cell proliferation will have ceased by approximately 8 weeks.
When spermatocytes are observed even in single seminiferous tubules, no proliferating Sertoli cells were found anywhere else in the same testis. In rodents, Sertoli cell replication has been found to cease upon entry of the first germ cell into meiosis [17]. Sertoli cell labelling was not observed in 3 of the 5 fowls at 8 weeks. This variation between individuals suggests that the commencement of meiosis may be a more practical indicator of the cessation of Sertoli cell replication in domestic fowl.
The volume density of the nucleus is usually proportional to the number of cells. However, the volume density of the Sertoli cells was higher than that of the germ cells at week 1 and equal at week 6. This shows that germ cells proliferate faster than Sertoli cells. The commencement of meioses depends on hormonal regulation, and the number of Sertoli cells germ cells is affected and altered by steroid hormones or endocrine disruptors [6,8]. The cessation of Sertoli cell replication with the commencement of meiosis may indicate that the former may depend on a general hormonal mechanism rather than local cell signalling. However the combined effect of Sertoli and germ cells on their development should not be underestimated. These results showed that Sertoli cell proliferation ceased when the volume densities of each type of cell nucleus converged. However, more study will be needed.


 XML Download
XML Download