

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Increased cardiovascular mortality is a serious problem in modern societies. Minimizing the risk of cardiac disease and alleviating the complications of cardiovascular dysfunction are the main therapeutic aims in modern medicine. Most heart diseases, regardless of etiology or pathogenesis, progress to congestive heart failure causing mortality [17]. The prognosis for patients with heart failure depends on the severity of cardiac dysfunction and the presence of complications and generally varies from guarded to poor. The single most powerful predictor for the development of heart failure is the presence of cardiac hypertrophy [18].
Cardiac concentric hypertrophy is a homeostatic response to elevated afterload that develops due to pressure overload (e.g. systemic hypertension or valvular stenosis). Although cardiac hypertrophy is the initial compensatory response to increased wall stress, it is followed by decompensating hypertrophy and ultimately leads to heart failure if the stimulus is sufficiently intense and prolonged. Cardiac concentric hypertrophy stimulated by an increase in blood pressure and wall stress is thought to be regulated by a number of intracellular signal transduction pathways including mitogen-activated protein kinase (MAPK), Janus kinase/signal transducers, activators of transcription (JAK/STAT), calcineurin, serine/threonine kinase, Akt and its downstream target, glycogen synthase kinase-3β [21]. Neurohormonal factors such as angiotensin II (Ang II) and endothelin-1 are activators of MAPK during conditions of pathological hypertrophy [27,29]. These observations have led to speculation that inhibition of these hypertrophic signals may provide effective clinical therapy for pathological hypertrophy and heart failure.
Based on considerable evidence accumulated during the last few years, much attention is focused on the use of naturally occurring botanicals for the prevention of many diseases. Epidemiological studies have shown a significant inverse association between dietary flavonoids and long-term mortality from coronary heart disease [5]. Epigallocatechin-3 gallate (EGCG), the major catechin derived from green tea, has been found to have protective effects on the cardiovascular system. These include anti-inflammatory effects [10], lowering of serum cholesterol levels, and reducing atherosclerosis [4]. The antioxidant properties of EGCG are well known [15] and have attracted considerable attention for the prevention of oxidative stress-related diseases such as ischemic heart diseases. The finding that reactive oxygen radicals significantly contribute to the genesis of reperfusion-induced dysrhythmias, contractile malfunction and vascular endothelium damage and that EGCG protects cardiac myocytes from ischemia/reperfusion injury also suggests that EGCG's cardioprotective effects may be mediated by free radical scavenging. However, there is little information about the effect of EGCG on cardiac hypertrophy. Recently, we demonstrated that EGCG inhibited platelet derived growth factor (PDGF)-BB-induced intracellular Ca2+ increase and extracellular signal-regulated kinase (ERK) activation in vascular smooth muscle cells (VSMCs) [1] and that EGCG could prevent Ang II-induced VSMCs hypertrophy through blocking c-Jun N-terminal kinases (JNKs) [29]. Moreover EGCG has been shown to inhibit PI3K/Akt and Erk1/2 pathways in several experimental models. These results suggest that EGCG could inhibit the signal pathways that regulate cardiac hypertrophy. Therefore, the purpose of this study is to investigate whether chronic administration of an oral daily dose of EGCG can attenuate cardiac hypertrophy induced by pressure overload in rats.
Materials and Methods
In vivo hypertrophy model
Male Sprague Dawley rats (7 weeks old, 200-220 g) were purchased from Koatech (Korea). The experimental protocol was approved by the Chungbuk National University Medical School Research Institutional Animal Care and Use Committee. All surgical procedures were performed on animals anesthetized with ketamine (80 mg/kg IP) and xylazine (5 mg/kg IP). Abdominal aortic constriction (AC) was performed using a 4-0 suture tied twice around the suprarenal aorta and a 21-gauge needle. The needle was then removed yielding a 0.8 mm internal diameter. Rats were randomly assigned to AC or sham-operated groups and the sham-operated rats underwent the same procedure, with the exception that the aorta was not constricted. A freshly prepared solution with different doses of EGCG (Sigma, USA) was supplied every day to aortic banded or sham-operated rats as the sole source of drinking water over a period of 21 consecutive days, whereas control animals were supplied with water from the same source, lacking EGCG (Fig. 1). Dietary administration was chosen to establish clinical relevance to human dietary habits. Establishment of cardiac hypertrophy was confirmed by echocardiography by measuring left ventricular (LV) wall thickness and dimensions, heart weight, and by histological analysis. The rats grew and gained weight during the course of the study, and as a control, we measured heart weight (HW) as a function of body weight (BW).
Hemodynamics measurements
Rat blood pressure was evaluated by direct cannulation of the right carotid and left femoral artery. Mean arterial blood pressure, heart rate and pressure gradient between carotid and femoral arterial pressure were obtained from a pressure transducer attached to each cannula, which was inserted through a fluid-filled catheter. The values were recorded using a computer data acquisition system (ML870; AD Instrument, Australia) after blood pressure was stable for 10 min.
Histological analysis and cardiomyocyte size measurement
All hearts were arrested in diastole with KCl (30 mM), followed by perfusion fixation with 10% paraformaldehyde. Fixed hearts were embedded in paraffin, and 4 mm thick sections were stained with hematoxylin and eosin for assessing overall morphology.
The surface area of a 2D silhouette of the myocyte was estimated by measurement of the length and width at 20 different randomly chosen points from a cross section of the LV free wall. Morphometric analysis was performed with isolution software (IMT, Korea). Our 2D surface area (length × width, µm2) is directly proportional to the surface area of a cylinder (2π × radius × length). The extent of LV fibrosis was measured using Cason's trichrome staining. Five sections of each heart were measured.
Echocardiography
After 21 days of aortic constriction, rats were anesthetized with intraperitoneal pentobarbital (50 mg/kg), and cardiac dimension and function were analyzed by 10-MHz pulsewave Doppler echocardiography (SONOACE 8800; Medison, Korea). Two dimensionally guided M-mode of LV at the papillary level was obtained from the parasternal long-axis view. For each rat, measurements were made from at least 4 beats. LV cavity dimension and wall thickness were measured, and percent change in LV dimension (fractional shortening; FS) and relative wall thickness (RWT) were calculated as follows: FS = [(LVDd-LVSd)/LVDd] × 100, where LVDd is LV dimension at end-diastole and LVSd is LV dimension at end-systole. RWT = (posterior wall thickness at end diastole)/ LVDd × 2.
Results
Effects of EGCG on load-induced increase in heart weight
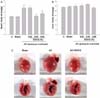
The amount of water intake was measured every day. Administration of EGCG in the drinking water at 0.02%, 0.04% and 0.08% did not alter water intake in either the sham-operated or aortic constricted rats. AC induced a 48% increase in HW/BW ratio compared to sham-operated rats. EGCG administration completely attenuated this increase (Fig. 2A). We found no significant difference between that the effect of 0.04% EGCG on pressure overload induced cardiac hypertrophy and that of 0.08% EGCG. The estimated EGCG intake of rats fed with 0.04% EGCG solution was 30 mg/kg. This estimate is based on an average consumption of 200 ml/day. These results are comparable with those in previous studies in which EGCG was non toxic to the liver and other organs after rats were injected intraperitoneally with 70-92 mg/kg daily [11]. Therefore, we selected 0.04% as the experimental dose. EGCG did not affect either body weight or liver weight (Fig. 2B). Next, we examined the effect of EGCG on HW/BW at various time intervals after AC surgery. During the 3 weeks after constriction, HW/BW ratio was significantly increased. The anti-hypertrophic effect of EGCG appeared as early as 1 week after surgery and was sustained during the course of the study (Fig. 4).
Effects of ECGC on cardiac and myocyte morphometric changes
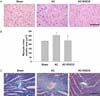
Aortic constriction caused a significant increase in both heart mass and LV wall thickness in comparison to sham-operated rats. AC-induced cardiac enlargement was almost completely prevented by EGCG, as shown in Fig. 2C. To examine whether EGCG changed heart size through the regulation of myocyte cell size, the myocyte cross-sectional area of the LV myocardium was measured in hearts from rats subjected to AC for 3 weeks (Fig. 3A & B). The cell area of aortic constricted rats was increased by 28% compared to sham-operated rats. This increase was almost completely abrogated by EGCG administration. AC also caused significant interstitial collagen deposition, as demonstrated by Masson's trichrome staining. This fibrosis was reduced in AC rats treated with EGCG for 3 weeks (Fig. 3C).
Effects of EGCG on hemodynamic loading conditions
Blockade of cardiac hypertrophy by EGCG in pressure-overloaded rats raised the possibility that EGCG might ameliorate the hypertension induced by AC, and thus the blockade of the hypertrophy might have been caused secondarily by reduction of afterload rather than by a direct cardiac effect. To rule out this possibility, blood pressure was measured. As expected, aortic constriction markedly increased the blood pressure and produced a large pressure gradient between the carotid and femoral arteries. However, EGCG did not affect the blood pressure, so that there was no significant difference in pressure gradient between aortic-banded rats and EGCG-treated aortic-banded rats (Table 1). Thus, EGCG attenuated load-induced cardiac hypertrophy in the presence of comparable hemodynamic loading conditions.
Effect of EGCG on cardiac function of aortic constricted rats
Eight weeks after constriction we examined the effect of EGCG on cardiac function using echocardiography (Fig. 5). Compared with sham-operated rats, aortic-banded rats displayed a substantial increase in interventricular septum thickness (IVSd) and LV posterior wall thickness in diastole (PWT). RWT was also markedly increased from 0.32 ± 0.04 in the sham group to 0.58 ± 0.05 in the AC group, which reveals evidence of concentric hypertrophy. Importantly, EGCG treatment significantly decreased PWT and RWT. EGCG-treated aortic-banded rats had a 20% increase in cardiac contractility, assessed by fractional shortening, compared to the AC group that did not receive EGCG (Table 2).
Discussion
In the present study, we found that a daily oral dose of the bioflavonoid EGCG can attenuate the cardiac hypertrophic changes induced by pressure overload. In addition, we showed that EGCG not only prevented cardiac hypertrophy resulting from pressure overload, but also improved cardiac performance mostly by reducing LV end diastolic and systolic dimensions.
Sustained high blood pressure is one of the strongest causes of the development of cardiac hypertrophy. EGCG and related bioflavonoids have shown to have vasodilatory effects probably induced both by facilitating diuretic vasorelaxation and by evoking endothelial-dependent vasorelaxation [14]. Likewise, it has been demonstrated that EGCG has an anti-hypertensive effect in the 5/6 nephrectomy rat model [20]. Therefore, blockage of pressure overload induced cardiac hypertrophy by EGCG raised the possibility that EGCG might ameliorate the hypertension induced by AC surgery and that the blockage of hypertrophy might have been secondary to afterload reduction rather than a direct cardiac effect. In this study, AC produced a large pressure gradient between carotid and femoral arteries; this gradient was not affected by EGCG treatment (Table 1). Hemodynamic measurements 3 weeks after surgery revealed that EGCG had no significant effect on the transstenotic aortic pressure gradients over time. By contrast, it has been recently reported that EGCG administration attenuated the increased blood pressure induced by aortic banding and essentially blocked the development of cardiac hypertrophy in this model [13]. The conflicting results from this study of Li et al. [13] are probably due to the difference in the dose of ECGC administered and the more severe banding conditions. Although the beneficial effect of EGCG on cardiovascular structure seems to be related to its blood pressure lowering effect, it is unlikely that the reduction in cardiac hypertrophy seen with this lower dose of EGCG was caused by relief of pressure overload. Our data suggests that the primary target organ of EGCG is the heart rather than blood vessels.
Previous reports that EGCG induced apoptosis of tumor cells raise the possibility that apoptosis in cardiomyocytes may occur in our animal model, leading to a reduction in cardiac mass. We did not perform the TUNEL assay in this study but it has been previously reported that pressure overload itself leads to an increase in TUNEL-positive cardiomyocytes [26]. Moreover, EGCG does not significantly affect the survival of cultured rat cardiomyocytes and has even been shown to have a protective effect against apoptosis induced by ischemic reperfusion injury [23]. Although EGCG has proapoptotic effects in transformed cells, apoptosis of cardiomyocytes does not appear to be responsible for the reduction of heart mass.
Pressure overload induces LV hypertrophy as a compensatory response to increased wall stress. This has been considered the central mechanism by which cardiac function is maintained within normal ranges in chronically overloaded hearts. Accordingly, suppression of myocardial hypertrophy is expected to cause heart failure. However, it has been suggested that under conditions of pressure overload, the development of cardiac hypertrophy and normalization of wall stress may not be a required compensatory response to pressure overload and may not be necessary to preserve cardiac function. Recently, it has been demonstrated that using genetically engineered mice that have markedly blunted growth responses to pressure overload, cardiac function was well maintained after loading (using a partial aortic constriction), despite the failure to correct wall stress. Indeed, function was in fact better maintained than in wild-type mice, in which hypertrophy ensued [6]. Moreover, Hill et al. [8] reported preserved cardiac output without hypertrophic compensation in the setting of pressure overload in thoracic aortic banding mice treated with cyclosporine A.
In this study, we examined the effect of EGCG on cardiac function using echocardiography. Treatment with EGCG greatly attenuated cardiac hypertrophy and there was no detectable impairment of cardiac function for 3 weeks. This result suggests that treatment with EGCG allowed the heart to adapt to pressure overload, even in the absence of LV hypertrophy.
Maintenance of cardiac output in the face of increased afterload without sufficient LV hypertrophy implies a positive inotropic effect. It has been known that under some experimental conditions, an increase in afterload causes an increase in ventricular inotropy. This homeometric autoregulation is called the Anrep effect [25]. Increased inotropy partially compensates for the increased end-systolic volume and decreased stroke volume caused by an increase in afterload. In this study, EGCG-treated aortic-banded rats had increased LV diastolic dimensions after 4 weeks. We presume that the Anrep effect in the EGCG-treated aortic-banded rat heart, which is under sustained pressure overload, was gradually exhausted, whereas cardiac function is still maintained. Additional work will be necessary to test this hypothesis and to evaluate the long-term effects of EGCG on cardiac function and survival in aortic constricted rats.
Reactive oxygen species (ROS) have been shown in a variety of cell types to act as intracellular signaling molecules in the stress response, leading to apoptosis, proliferation, and transformation. In cardiomyocytes, ROS have been found to be involved in cardiac hypertrophy in vitro [22] and to mediate hypertrophy induced by several stimuli, such as mechanical stretch [19], Ang II [16], and tumor necrosis factor-α [7]. EGCG has been known to have potent antioxidant activity [15]. We found that the levels of malondialdehyde (MDA) were markedly increased in the AC group and EGCG treatment resulted in an almost complete elimination of the increase in MDA (data not shown). Therefore, we propose that the antioxidant effect might also play a role in the prevention of cardiac hypertrophy in AC rats treated with EGCG.
A different location of the aortic constriction may differently activate the signal transduction pathway. When the aortic arch was constricted, mechanical force may have been the major cause of cardiac hypertrophy. In contrast, the renin-angiotensin system (RAS) may be involved predominantly in suprarenal abdominal aorta constriction and contribute to the initial development of cardiac hypertrophy and sympathetic activation in the compensated heart. It has been shown that losartan, an Ang II AT1 receptor antagonist, attenuates cardiac hypertrophy in suprarenal abdominal aorta constricted rat [12], but has no effect on the weight gain of the ventricle during aortic arch constriction [2]. Given that EGCG can inhibit Ang II mediated signal transduction in cultured cardiomyocytes [28], the antihypertrophic effect of EGCG in the present study could have been mediated by RAS blockage. However, EGCG had no significant effect on systemic blood pressure, which sensitive to Ang II. These findings suggest that inhibition of RAS by EGCG does not play a major role in the EGCG-mediated antihypertrophic effect. Calcineurin inhibitors, such as cyclosporine A and FK506 have been well known to inhibit cardiac hypertrophy. However, nephrotoxicity and the immunosuppressive effect of calcineurin inhibitors limit their therapeutic benefit [24]. In our experiment, EGCG completely inhibited cardiac hypertrophy induced by pressure overload without significant effects on normal heart and liver. There was no evidence of nonspecific toxicity since no effects on normal growth, weight gain, or physical activity were found. Therefore, EGCG, or its derivatives, might be a useful modality to suppress cardiac hypertrophy.
In congestive heart failure by concentric hypertrophy, positive inotropic agents may significantly reduce intraventricular volume and cardiac output and thus contribute to a increase in mortality, so positive inotropes are prohibited in the treatment of concentric hypertrophy. Hotta et al. [9] showed that EGCG (10-6, 10-5 M) increased LV developed pressure and claimed that EGCG represents a positive inotropic effect via the nitric oxide pathway [9]. Given the low bioavailability of EGCG, it is unlikely that EGCG had an inotropic effect in our experimental system.
In this study, we demonstrated that EGCG (approximately 30 mg EGCG/kg body weight) prevents the development of cardiac hypertrophy induced by pressure overload. It has been reported that most of the EGCG does not get into the blood, but is excreted through the bile to the colon, so that serum concentration of total EGCG was 43 nM in male rats treated with 75 mg EGCG/kg body weight by gavage [3]. Even if such a low dose of EGCG had a positive inotropic effect, this effect partially compensated for the increase in afterload and helped the heart to adapt to pressure overload at the development of hypertrophy stage. However, we can not exclude the possibility that EGCG would have a harmful effect on advanced chronic heart failure and a severely dilated ventricle. Future studies are necessary to examine whether EGCG can regress established cardiac hypertrophy, which might be of greater clinical significance because the patient would most likely already have hypertrophy at the initiation of treatment.
In conclusion, this study demonstrates that EGCG completely inhibited cardiac hypertrophy and improved cardiac performance in pressure overloaded hearts. These findings may have important clinical implications in developing new therapeutic strategies to prevent the transition from cardiac hypertrophy to heart failure.





 XML Download
XML Download