Introduction
Infectious salmon anemia virus (ISAV), an economically important pathogen of farmed Atlantic salmon in the northern hemisphere, is a type species under the new genus Isavirus within the family Orthomyxoviridae [1,21,42]. Like the influenza A and B viruses, ISAV particles are enveloped with a segmented genome comprising eight single-stranded RNA segments of negative polarity [3,21,28]. As for the influenza viruses, ISAV has surface projections that exhibit hemagglutination, receptor destroying and fusion activities [1,13,21~23,27]. Since its first description on the Norwegian salmon farms in 1984, the disease or presence of the virus has been reported in aquaculture facilities of several other countries including Canada [1,21,41,42]. Although ISAV causes clinical disease principally in farmed Atlantic salmon [21,33,34,35,41], the virus also infects and replicates in wild fish that may become asymptomatic, probably life-long carriers of the virus [1,6,21,33~35,38,41,44,45]. It is widely believed that virulent ISAV strains emerge from these background-covert infections in wild fishery, become adapted to aquaculture practices, and cause clinical disease in marine-farmed Atlantic salmon [16,21,33~35]. ISAV strains vary in their antigenicity, pathogenicity, and ability to replicate and/or cause cytopathic effects (CPE) in the available fish cell lines [6,16,21,23,30,41].
Virus isolation and neutralization in cell culture, conventional reverse transcription-polymerase chain reaction (RT-PCR), and indirect fluorescent antibody tests are the methods commonly being used to detect ISAV in biological samples [2,4,7~12,17~21,24,26,29,30,36,41,42,45]. An in situ hybridization method for ISAV detection has also been described [12,29].
Several cell lines including primary cells, ASK-2, SHK-1, CHSE-214, TO, AS and Rtgill-W1 have been used to isolate ISAV from field samples [4,11,18~21,30,41~43,46]. Primary cell culture and AS and Rtgill-W1 cells are rarely used for ISAV isolation; primary cells show variable susceptibility to ISAV and give low virus yield while hemadsorption or an indirect fluorescent antibody test is required to detect the replication of ISAV in AS and Rtgill-W1 cells because the virus replicates poorly without causing any CPE in these cell lines [4,21]. Although SHK-1, TO, ASK-2 and CHSE-214 are commonly employed cell lines for ISAV isolation, these cell lines also have limitations [11,18~21,30,41,43,46]. The former three cell lines require very low split ratios and the latter cell line does not support the growth of all ISAV isolates. CPE development may take up to 17 days in CHSE-214 cells with a virus yield lower than SHK-1. SHK-1 is a very delicate cell line requiring complex growth medium and sometimes, it loses sensitivity at higher passages [21]; moreover, some ISAV isolates produce poorly defined and slowly developing CPE in the SHK-1 cell line [21]. Recovering the virus from some ISA outbreaks or RT-PCR positive samples in ASK-2, TO, and SHK-1 cell lines has not been possible. The availability of the TO cell line is also at the discretion of the patent holder. Virus typing by neutralization test in macrophage-like (SHK-1 and TO) cell lines is also considered unreliable because of the serum toxicity or enhancement of ISAV infection of these cell lines by ISAV-specific antibodies via an Fc-receptor mediated endocytosis [15,21]. Indirect fluorescent antibody testing and conventional RT-PCR including its variants (such as nested RT-PCR and RT-PCR-coupled denaturing gradient gel electrophoresis) are available only in yes and no formats [2,7~9,12,17,24,26, 29,30,36,42]. The former test, like in situ hybridization, is very subjective and later depends upon end-point detection and carries a high risk of contamination. In general, all the above methods are time-consuming, laborious and require several hours to several days for their completion with a low throughput.
We have previously described the development of a single-tube real-time RT-PCR assay for the quantitative detection of ISAV load in biological samples using SYBR Green chemistry and LightCycler technology [30] to counter the limitations of the above-mentioned methods. The assay utilized a F5/R5 primer set targeting ISAV RNA segment 8. An r2 value of 96.2%, a slope of -3.771, and r values of 0.98 and 0.96 for precision- within and -between assays respectively suggested that the assay was highly reproducible and that there was a strong linear association between viral RNA levels and threshold cycle (Ct) values. The assay was found to be 100 times more sensitive than the conventional RT-PCR for ISAV and could be completed within 80 min.
The purpose of the present study was twofold: 1) to demonstrate that the optimized real-time RT-PCR conditions as described previously [30] could detect ISAV isolates of different geographic origins, and 2) to analyze the growth patterns and replication of selected ISAV isolates in CHSE-214 cells in order to characterize their CHSE-phenotypes.
Materials and Methods
Experimental design
This study was carried out in three phases. In Phase I, the ability of ISAV isolates from different geographic origins and/or laboratory sources to cause CPE in CHSE-214 cells was evaluated. For this purpose, each of the selected isolates was inoculated into a CHSE-monolayer and the monolayer was observed for the appearance of CPE for up to 14 days post-infection. To confirm the presence of ISAV isolates in biological samples that did not cause CPE in CHSE-214 cells, each ISAV isolate was also grown in TO cells. In Phase II, the ability of optimized real-time RT-PCR assay conditions to detect ISAV isolates of different geographic origins or laboratory sources was tested. For this purpose, the total RNA from virus infected tissue culture suspensions or infected samples was extracted and amplified using SYBR Green real-time RT-PCR. In Phase III, the replication of the following seven ISAV isolates in CHSE-214 cells were analyzed by CPE appearance, real-time RT-PCR and viral titrations: NBISA01, U5575-1, Scottish 1490/98, HKS-36, RPC/NB 01 0090-2, RPC/NB 02 1223-4, and RPC/NB 00 1016-1. NBISA01, U5575-1, and HKS-36 served as positive controls in this experiment for replicating cytopathic, non-replicating non-cytopathic, and replicating non-cytopathic CHSE-phenotypes of ISAV, respectively, because we knew of their CHSE-phenotypes from our previous studies [18~20,30]. For testing the replication pattern of each of these isolates in CHSE-214 cells, several CHSE-214 cell monolayers were grown. Each isolate was inoculated into nine CHSE-monolayers, which were then incubated at 16℃. CHSE-monolayers inoculated with phosphate buffered saline (PBS) were also included as controls. Three of the tissue culture infected flasks for each isolate were removed from incubation at days zero (immediately upon adding maintenance medium post-inoculation), 7 and 14 post-inoculation, observed for CPE and frozen at -80℃. At the end of all incubations, the contents from infected as well as uninfected control tissue culture flasks were harvested and their total RNA was extracted and amplified by SYBR Green real-time RT-PCR. Similarly, each of these isolates was inoculated into another set of three CHSE-monolayers and viral titrations of infected CHSE-214 cell contents harvested at post-infection day zero, 7, and 14 were performed on TO cells in 48-well tissue culture plates. The viral RNA at different days post-infection in these infected cell harvests was also analyzed by SYBR Green real-time RT-PCR assay in order to find the corresponding Ct value.
Cell culture and viral isolates
A total of 24 (17 North American and seven European) ISAV isolates were used in this study (Table 1). The criteria used for the selection of ISAV isolates were their geographic origin and laboratory source.
Two cell lines, TO and CHSE-214, were used to propagate ISAV in this study. For infection, CHSE-214 cell monolayers were grown at 16℃ in Hank's minimum essential medium (Invitrogen, USA) supplemented with 10% bovine fetal serum (FBS), 100 µg/ml streptomycin, 0.25 µg/ml amphotericin B, and 100 U/ml penicillin in 25 cm2 tissue culture flask(s) as previously reported [18,19,30]. When approximately 85% confluence was attained, the monolayer(s) were washed with PBS, inoculated with virus, and incubated at room temperature for 1 h to allow for virus adsorption. The maintenance medium (FBS reduced to 5%) was then added and the flasks were incubated at 16℃ for up to 14 days. The tissue culture virus suspension was harvested, aliquots dispensed and stored at -80℃ for further use. TO cells were grown as described previously [30,47]. Briefly, TO cell monolayers were grown at room temperature in Eagle's minimum essential medium containing Hanks' salts (BioWhittaker, USA) supplemented with 10% heat inactivated FBS, 292 µg/ml L-glutamine (Invitrogen, USA), 200 µg/ml gentamicin (Sigma, USA) and 1% non-essential amino acids (Sigma, USA). Similarly, TO cell monolayers were inoculated with virus and the infected flasks were incubated at 16℃ for up to 10 days. ISAV isolates were purposely handled on separate days to minimize the risk of cross-contamination.
RNA extraction
The total RNA from virus-infected cell culture suspensions, infected samples or uninfected control samples was extracted using TRIZOL reagent following the manufacturer's instructions (Invitrogen, USA) and as described previously [19,30] with minor modifications. The starting sample volume for all RNA extraction was 300 µl of virus-infected (or uninfected) tissue culture suspensions. Triplicate RNA extractions for each sample were performed. The RNA extractions for each ISAV isolate were carried out separately on different days in order to minimize contamination.
Viral RNA amplifications
All real-time RT-PCR reactions for ISAV with the F5/R5 primer pair targeting ISAV RNA segment 8 were carried out using RNA Amplification SYBR Green I (Roche, Canada) as described previously [30]. The 20 µl reaction volume for real-time RT-PCR contained 4.0 µl of 5 × RT-PCR reaction mix SYBR Green I, 3.0 µl of 5 × resolution solution, 5.0 mM of MgCl2, 0.3 µM of forward primer-F5 (5' GAA GAG TCA GGA TGC CAA GAC G-3') and equal concentrations of the reverse primer-R5 (5' GAA GTC GAT GAT CTG CAG CGA-3'), 9.4 µl of PCR-grade water, 0.4 µl of enzyme mix, and 1.0 µl of template (≥0.632 ng/µl). All real-time reactions were run in the LightCycler (Roche, Canada) with the following thermal profile: a single cycle of reverse transcription for 30 min at 55℃ and a pre-denaturation step for 30 sec at 95℃ followed by 50 amplification cycles each consisting of denaturation for 5 sec at 95℃, annealing for 10 sec at 59℃, and elongation for 10 sec at 72℃. An additional step of 80℃ for the fluorescence acquisition was also included at the end of each amplification cycle to enhance the specificity of the fluorescent signal. This step was added because our previous studies [30] indicated that the melting temperatures (Tm) for ISAV-specific PCR products were higher than 80℃, whereas nearly all non-specific products melt away below this temperature. After amplification was concluded, a melting curve analysis was performed by slowly heating the cooled reaction products from 70℃ to 95℃ in 0.1℃/s increments to confirm the specificity of viral amplicon(s). To further confirm the specificity of the amplification products, the amplified PCR products of some reactions were also resolved by gel electrophoresis. The products were resolved by 1% agarose gel electrophoresis in 0.5 × TBE buffer, visualized by staining with ethidium bromide, and photographed under 304 nm UV light. A sample was considered positive if it was positive by the melting curve, the fluorescent signal was above the average background, and the Ct value was ≤40. All the samples showing Ct values ≥40 were considered negative provided they are were also negative by the melting curve.
Data analyses
The amplification plots and melting curve data for all real-time RT-PCR runs were analyzed using the LightCycler software version 3.5 (Roche, Canada). One way analysis of variance (ANOVA) at 1% α level was used to compare the mean Ct values of viral RNA levels for each ISAV isolate in CHSE-214 cells at zero, 7, and 14 days post-inoculation. The ANOVA was performed using Minitab 13 software (Minitab, USA).
Results
Detection of ISAV isolates of different geographic origins by SYBR Green real-time RT-PCR
Twenty-four North American and European ISAV isolates tested in this study are shown in Table 1. The results indicate that the SYBR Green real-time RT-PCR assay could detect all the ISAV isolates selected for this study. The melting curve and gel analyses (not shown) indicated that the amplification products were virus-specific as single melting peaks characteristic of the viral amplicon and band sizes of approximately 220 bp were obtained for all the isolates tested. Gel electrophoresis of the amplification reaction products that had Ct values equal or greater than 36 did not yield virus-specific bands. The melting temperatures (Tm) for different viral isolate-specific amplification products ranged from 82.32 to 84.05℃. A difference of 0.3 to 1.5℃ in the Tm(s) was observed for some ISAV isolates between runs.
Differentiation between CHSE-positive and -negative phenotypes of ISAV
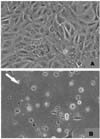
The ISAV isolates were also tested for their ability to cause CPE in CHSE-214 cells and TO cells (Fig. 1 and 2; Tables 1 and 2). The results indicate that ISAV isolates, based on the absence or presence of CPE in CHSE-214 cells could be divided into two CHSE-phenotypes: CHSE-positive and CHSE-negative. The CHSE-positive ISAV phenotype caused CPE in CHSE-214 cells; the CPE involved only a limited number of cells in the monolayer and appeared as localized clusters of cell rounding and lyses (Fig. 1). The CHSE-negative ISAV phenotype, however, did not induce CPE in these cells. These results also indicate that both CHSE-positive and negative ISAV phenotypes are present in Canada whereas nearly all European ISAV isolates belong to a CHSE-negative phenotype. All ISAV isolates caused CPE in the TO cells (Fig. 2).
Characterization of three ISAV CHSE-phenotypes
The replication of the following seven ISAV isolates in CHSE-214 cells was analyzed by CPE appearance, viral RNA quantitation by SYBR Green real-time RT-PCR, and viral titrations of the infected CHSE cell contents in TO cells: NBISA01, U5575-1, Scottish 1490/98, HKS-36, RPC/NB 01-0090-2, RPC/NB 02-1223-4, and RPC/NB 00-1016-1 (Tables 1 and 2). The results suggest that of the eight ISAV isolates, only NBISA01 caused CPE in CHSE-214; the CPE was first observed on day 7 post-infection. All other isolates, including, HKS-36, did not cause CPE in CHSE-214 cells throughout the 14 days of incubation (Table 2). The temporal analysis of viral RNA in CHSE-214 cells at post-infection days zero, 7, and 14, however, suggests that both HKS-36 and NBISA01 replicated in the CHSE-214 cell line as there was a significant difference (p < 0.01) between the Ct values for viral RNA levels of these two isolates at different post-infection days (Table 2). The data further suggest that the ISAV isolates U5575-1, 1490/98, RPC/NB 01-0090-2, RPC/NB 02-1223-4, and RPC/NB 00-1016-1 did not replicate in CHSE-214 cells as differences between the Ct values for viral RNA levels of these isolates at different post-infection days were not significant (p > 0.01) (Table 2). The amplification plots and melting temperatures of the three representative isolates are shown in Figures 3 and 4. Melting curve analyses confirmed that real-time RT-PCR products were virus-specific as Tm(s) for amplification products were between 83 and 84.5℃. The viral titers and corresponding Ct values of the infected CHSE cell contents harvested at post-infection days zero, 7, and 14 are presented in Table 2. The viral titers of infected CHSE harvests at different post-infection days further confirm that only the two ISAV isolates, NBISA01 and HKS-36, replicated in CHSE-214 cells. These results also suggest that ISAV isolates could be grouped into three CHSE-phenotypes: replicating cytopathic (causes CPE and replicates), replicating non-cytopathic (replicates without causing any CPE or cell death), and non-replicating non-cytopathic (does not replicate or cause CPE).
Discussion
The results of this study verify that the SYBR Green real-time RT-PCR is capable of detecting ISAV isolates of different geographic origins. Moreover, just by the presence or absence of CPE in CHSE-214 cells, ISAV isolates could be divided into two CHSE-phenotypes. To distinguish replicating non-cytopathic ISAV strains from non-replicating non-cytopathic strains, viral quantification at different post-infection times could be used to discriminate between the two possibilities. The SYBR Green real-time RT-PCR is a highly effective tool for this purpose as we have demonstrated that the assay could differentiate between replicating noncytopathic ISAV strains and non-replicating non-cytopathic ISAV strains with greater sensitivity than isolation of the virus from the CHSE-214 cells. Thus, this study demonstrated the usefulness of the SYBR Green real-time RT-PCR for the detection and quantification as well as study of the epidemiology and pathogenesis of ISAV strains [25,30~32,48].
SYBR Green 1 and the hydrolysis probe are the two most widely used real-time chemistries for signal generation [31,32,48]. The SYBR Green 1-based assays have two important advantages over the hydrolysis probe-based assays: 1) they are simpler and less expensive, and 2) they allow melting curve analysis at the end of amplification reactions, which is used to confirm the specificity of the reaction products. SYBR Green 1 was the chemistry used for fluorescent signal generation in all the real-time RT-PCR reactions performed in this study. Although the gel analyses of amplification reactions for most of the North American and European ISAV isolates confirmed that amplification products were virus-specific, as bands of approximately 220 bp sizes were obtained following gel electrophoresis and ethidium bromide staining, the gel analyses of few amplification reactions that had Ct values equal or greater than 36 and were positive by melting curve did not yield any virus-specific bands. This observation suggests that the real-time detection format is more sensitive than is gel electrophoresis of PCR products. This observation is in agreement with previous reports that suggest the higher sensitivity of the real-time format over agarose gels [24,25,30]. We have reported previously that agarose gels have a limited dynamic range [30]. The high Ct values for some ISAV isolates imply that these isolates are either from carrier fish or they represent a slowly replicating non-pathogenic ISAV strain(s) [37]. Hodneland and Endresen [14] have recently reported similar observations with Salmonid alphavirus-infected fish and suggested that higher Ct values might reflect pre-viraemic, post-viraemic or carrier fish status where the host is in the process of clearing the virus. The Tm(s) for different viral isolate-specific amplification products ranged from 82.32 to 84.05℃ and a difference of 0.3 to 1.0℃ in the Tm(s) was observed for some ISAV isolates between the runs. Whether this variation in Tm is due to nucleotide differences or pipeting errors is not known. The latter is more likely because of the following two reasons: 1) a variety of factors including buffer conditions and stability of the SYBR Green dye may affect the Tm values [25]; and 2) a substantial nucleotide change is required to cause a shift in the Tm(s) of the amplicon while using SYBR Green chemistry since the DNA binding dyes (SYBR Green 1) probably increase the melting transition of the amplicon [25]. The results that of the seven ISAV isolates whose replication and ability to cause CPE in the CHSE-214 cells analyzed, only NBISA01 caused CPE, U5575-1 did not replicate and cause CPE, and HKS-36 replicated without inducing any CPE in the cells, are consistent with our previous reports [18~20,30].
The identification of three CHSE-phenotypes of ISAV in North America may have important implications from diagnostic and biological points of views. For example, fish positive by conventional RT-PCR does not imply that the relevant ISAV isolate is a replicating strain and vice versa; the virulence or pathogenicity indices of such isolates should always be considered and tested. It does imply, however, that the field samples negative for ISAV by CHSE-214 cell culture (absence of ISAV-characteristic CPE) are not necessarily negative as they may contain the non-cytopathic replicating ISAV phenotype. Similar findings for other viral pathogens have also been reported. Thus, three bovine viral diarrhea virus (BVDV) biotypes have been described: the non-cytopathic biotype does not cause any CPE in cultured epithelial and lymphoid cells, the cytopathic biotype causes CPE in both epithelial and lymphoid cells, and the lymphopathic biotype causes CPE but only in lymphoid cells [40]. It has also been suggested that the lymphopathic BVDV strains correlate with high virulence in vivo [5,40] and it is widely believed that cytopathic BVDV strains arise by mutation of non-cytopathic strains [5]. Whether a similar correlation between the in vivo ISAV virulence and its CHSE-phenotypes exists remains to be investigated. The exact reason why some ISAV isolates cause CPE and others do not in the CHSE-214 cell line remains unknown.
Optimized SYBR Green real-time RT-PCR conditions are flexible in detecting ISAV isolates of different geographic origins. This assay, along with the capacity of ISAV isolates to replicate and cause CPE in the CHSE-214 cell line, could be used to group ISAV isolates into one of the three CHSE phenotypes: non-replicating, replicating non-cytopathic, and replicating cytopathic. Further, this assay could potentially be used in studying the neutralization of non-cytopathic replicating ISAV strains by specific antiserum in the CHSE-214 cells and thereby have the potential to assist in serotyping non-cytopathic replicating ISAV strains that do not cause CPE in one or more cell lines. The results of this study help to explain the failure to isolate ISAV in the CHSE-214 cell line from some ISA positive fish or RT-PCR positive samples.