

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
T-cell internal antigen-1 (TIA-1) and TIA-1-related protein (TIAR) are two closely related RNA-binding proteins that regulate gene expression by binding to uridine-rich RNA sites. Endogenous and recombinant wild-type TIA-1 is distributed both in the nucleus and cytoplasm [15]. The role of TIA-1 varies, depending on its subcellular localization. In the nucleus, TIA-1 regulates alternative pre-mRNA splicing [728]. In the cytoplasm, TIA-1 regulates mRNA translation by binding to AU-rich elements in the 3′-untranslated region (3′UTR) of mRNA [1120]. In addition, when cells are exposed to environmental stressors such as oxidative stress, heat shock, or virus infection, TIA-1 shuttles from the nucleus to the cytoplasm to sequester untranslated mRNA at cytoplasmic foci called stress granules (SGs) [15]. Subsequently, cells enter a relatively stable state by recruiting mRNA transcripts into SGs, thereby inhibiting protein translation in order to adapt to the altered condition. The assembly of SGs is initiated by the phosphorylation of eukaryotic initiation factor (eIF)2α, which is a component of the eIF2-GTP-tRNAMet ternary complex that associates with the 40S complex and scans for an initiation codon. The phosphorylation of eIF2α leads to a decrease in abundance of the eIF2 ternary complex [13]. RNA-binding protein TIA-1 and TIAR sequesters untranslated mRNAs at SGs in the cytoplasm [15].
Canonical SGs contain some specific components. The core constituents of SGs are aggregates of stalled 48S translation pre-initiation complexes that include small ribosomal subunits and eIF3, eIF4E, and eIF4G [12]. In addition, SGs also contain RNA-binding proteins such as poly(A)-binding protein (PABP) 1, Ras-GTPase-activating protein-binding protein (G3BP), and human antigen R (HuR) [81525]. The assembly of SGs involves a highly dynamic process. The SG components TIA-1 and PABP-I continuously shuttle in and out of SGs [14]. Moreover, the accumulation of untranslated mRNA at SGs is also dynamic because drugs that stabilize (e.g., cycloheximide [CHX]) or destabilize (e.g., puromycin) polysomes inhibit or promote SG assembly, respectively [14].
TIA-1 is an RNA-binding protein that possesses three amino-terminal RNA recognition motifs (RRMs) and a carboxy-terminal prion-related domain (PRD), which are both necessary for SG assembly. The RRM is required to recruit RNA, while the PRD is required to aggregate RNA, RNA-binding protein, and other SG components to form SGs [10]. Full-length TIA-1 expression induces the assembly of SGs. Overexpression of PRD results in cytoplasmic aggregates that sequester endogenous TIA-1 [15].
SGs were first observed in the cytoplasm of tomato cells that had been exposed to heat shock [18]. Similar SG structures were detected in mammalian cells that were subjected to various environmental stresses. Moreover, TIA-1 has been identified as a key component in mammalian SGs [15]. These reports indicate that SG is an evolutionally conserved response to environmental stress. Previous reports have provided clues on SG formation in chickens [31617]. For instance, heat-shock protein (hsp)24 is recruited to distinct cytoplasmic insoluble aggregates, which are probably SGs [3]. An in vivo experiment showed that SGs are present in the cochlear cells of chickens that had been treated with the ototoxic antibiotic, gentamicin [17]. However, the detailed features of SG formation in chicken cells have not been described. In particular, the role of TIA-1 in chicken SG assembly has not been elucidated, although TIA-1 and TIAR have been proven to be necessary for chicken DT-40 cell viability [16]. Herein, we report that the exogenous expression of chicken TIA-1 (cTIA-1) facilitates the assembly of classical SGs in both human and chicken cells. In addition, the overexpression of chicken PRD (cPRD) bearing an amino-terminal green fluorescent protein (GFP) tag (pntGFP-cPRD) or a Flag tag (pFlag-cPRD) but not a carboxyl-terminal GFP tag (pctGFP-cPRD) induces the formation of typical SGs, indicating that the GFP tag affects cPRD localization. Finally, endogenous cTIA-1 has been shown to be recruited to SGs in chicken cells and tissues under environmental stress.
Materials and Methods
Reagents and antibodies
Mouse monoclonal anti-Flag (F1804), rabbit polyclonal anti-GFP (G1544), sodium arsenite (ARS, S7400), and CHX (C7698) were purchased from Sigma-Aldrich (USA). Goat polyclonal anti-TIA-1 (sc-1751) and mouse monoclonal anti-PABP (sc-32318) antibodies were purchased from Santa Cruz (USA). Mouse monoclonal anti-G3BP (ab56574) antibody was purchased from Abcam (USA). Rabbit polyclonal anti-eIF3A (sc-2538) antibody was purchased from Cell Signaling Technology (USA). Horseradish peroxidase-conjugated donkey anti-goat, goat anti-rabbit, and anti-mouse secondary antibodies were purchased from Jackson Immuno Research (USA). Alexa Fluor 488-labeled donkey anti-goat IgG (A11055), Alexa Fluor 594-labeled goat anti-rabbit (A11012), and goat anti-mouse (A-11005) were purchased from Thermo Fisher Scientific (USA).
Cells and viruses
HeLa and DF-1 chicken fibroblast cells were purchased from American Type Culture Collection (ATCC; USA). Chicken embryo fibroblast (CEF) primary cells were prepared from 9-day-old specific-pathogen-free (SPF) embryonated chicken eggs as described previously [26]. Cells were maintained in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (FBS; Gibco, USA). Newcastle disease virus (NDV) strain Herts/33 was obtained from the China Institute of Veterinary Drug Control (China).
Cloning and plasmids
Full-length cTIA-1 was amplified from cDNA isolated from CEFs by using Phusion High-Fidelity DNA polymerase (New England Biolabs, USA). Expression plasmids of C-terminal-GFP-tagged cTIA-1 full-length gene (cTIA-1 full), RRMs (cTIA-1 RRM), and PRD (cTIA-1 PRD) were constructed by inserting the full-length gene, the first 654 nucleotides, or the last 465 nucleotides of cTIA-1, respectively, into the EcoRI/SalI sites of plasmid pEGFP-N1 (Clontech, USA). Expression plasmids of Flag-tagged cTIA-1 full, RRM, and PRD were constructed by inserting the corresponding sequence into the EcoRI/BamHI sites of plasmid p3XFLAG-CMV-14 (Sigma-Aldrich). The expression plasmids of N-terminal-GFP-tagged chicken TIA-1 PRD were constructed by inserting the last 465 nucleotides of cTIA-1 into the EcoRI/BamHI sites of plasmid GFP-C3 (Clontech) (Fig. 1).
Stress treatment and pharmacological experiments
For oxidative stress, cells were exposed to 0.5–1 mM ARS in complete medium for 30 min at 37℃. For heat shock, HeLa and DF-1 cells were cultured in complete medium for 40 min at 42℃ and 47℃, respectively. For virus infection, the HeLa cells were infected with NDV at a multiplicity of infection of 1 at 37℃. Following a 1-h absorption period, unattached viruses were removed, and the cells washed three times with phosphate-buffered saline and cultured in a maintenance media at 37℃. Cells were harvested at 18 h post-infection.
For pharmacological experiments, cells were treated with CHX (100 µg/mL), together with vehicle controls for 2 h before ARS treatment. Subsequently, the cells were subjected to immunofluorescence (IF) at the indicated times post-infection.
Immunofluorescence microscopy
HeLa cells grown on glass coverslips were transfected with GFP-tagged TIA-1 full, RRMs, or PRD. Approximately 24 h after transfection, the cells were subjected to various stressors. At indicated times post-treatment, the cells were fixed in 4% neutral formaldehyde and visualized by using a Nikon C1-si confocal fluorescence microscope (Nikon Instruments, Japan). For the colocalization assay, after fixation, the cells were permeabilized with 1× Tris-buffered saline with Tween-20 (TBST) with 0.5% Triton X-100 for 10 min, incubated in a blocking buffer (1× TBST with 3% bovine serum albumin), and then incubated with primary antibodies for 1 h at 37℃. The cells were washed three times with 1× TBST and incubated with secondary antibodies conjugated to Alexa Fluor 594 (Life Technologies, USA). The cells were then washed three times with 1× TBST. Nuclei were stained with 4′, 6-diamidino-2-phenylindole (DAPI; Beyotime Biotech, China) at 37℃ for 10 min. After washing four times with 1× TBST, the coverslips were mounted on slide glasses and visualized.
Animal experiments
SPF chicken embryos were obtained from Merial (Merial Vital Laboratory Animal Technology, China) and incubated in a thermotank. After hatching, the chickens were housed in cages under biosafety conditions and a 12-h light/dark cycle with free access to water and food. Care and maintenance of all animals were in accordance with the Institutional Animal Care and Use Committee guidelines established by the Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Science (permit No. SHVRI-Po-2016070686). Six 7-day- old SPF chickens were randomly divided into two groups of 3 and mock treated or treated with heat shock (40℃ ± 1℃ for 10 h). The mRNA level of the TIA-1 gene in the tissue was measured by performing real-time quantitative reverse transcription-polymerase chain reaction (qRT-PCR) as previously described [23]. Chicken hearts were immediately dissected and sliced by using a freezing microtome (CM1950; Leica, Germany). The IF assay was subsequently performed by using goat polyclonal anti-TIA-1 antibody as the primary antibody, as described earlier [24].
Results
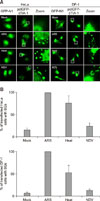
Ectopic and exogenous expression of cTIA-1 induces the formation of cytoplasmic granules in HeLa and DF-1 cells under various stressors
The formation of SGs was characterized by assessing the fluorescent spots of cytoplasmic TIA-1 protein under various stressors such as oxidative stress, heat shock, and virus infection. We constructed the pctGFP-cTIA-1 and transfected it into HeLa and DF-1 cells. The results showed that pctGFP-cTIA-1 accumulated to form distinct dots in the cytoplasm in both ARS- and heat-shock-treated HeLa cells. The TIA-1 dots were also detected in NDV-infected HeLa cells (panel A in Fig. 2; columns 2 and 3). In comparison, the transfection of the pEGFP-N1 vector, as a negative control (mock), resulted in a diffuse distribution of green fluorescence in whole cells (panel A in Fig. 2; column 1). Similar TIA-1 dots were observed in the cytoplasm of DF-1 cells (panel A in Fig. 2; right panel). The TIA-1 dots were quantified visually by manual counting. Approximately 10 pictures in 20 high-powered fields were randomly captured at different locations. Panel B in Fig. 2 shows that with ARS treatment, TIA-1 dots were observed in 100% of pctGFP-cTIA-1 transfected HeLa cells. Approximately 75% and 25% of the pctGFP-cTIA-1 transfected cells apparently contained cytoplasmic granules in heat-shock-treated and virus-infected HeLa cells, respectively (panel B in Fig. 2; left panel). Similar trends were observed in pctGFP-cTIA-1 transfected DF-1 cells under the same stress conditions (panel B in Fig. 2; right panel). Overexpression of pctGFP-cTIA-1 induced the production of a few TIA-1 dots in pctGFP-cTIA-1 transfected cells (~20% for HeLa and ~10% for DF-1 cells) in the absence of treatment, thereby indicating that the ectopic expression of cTIA-1 alone induces the formation of cytoplasmic granules.
Ectopic expression of chicken TIA-1 induces bona fide SGs in HeLa cells
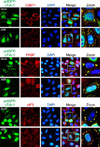
Because the transfection of pctGFP-cTIA-1 induced the formation of TIA-1 dots in HeLa and DF-1 cells, we then evaluated whether these dots were actual SG dots. G3BP, PABP, and eIF3 are considered authentic SG markers under various stressors [121525], and pctGFP-cTIA-1 colocalization with those endogenous SG markers was investigated to confirm whether the observed TIA-1 dots were actual SGs. Fig. 3 shows the colocalization of pctGFP-cTIA-1 and endogenous G3BP1, PABP, and eIF3 in ARS-treated cells. Moreover, in some mock-treated cells, the transfection of pctGFP-cTIA-1 induced the formation of cytoplasmic dots, which also colocalized with endogenous G3BP1, PABP, and eIF3, respectively. These results suggest that the overexpression of pctGFP-cTIA-1 induces the formation of SG dots in cells, regardless of the presence of stressors.
pctGFP-cPRD induces the formation of large granules that do not disassemble after CHX treatment
Previous reports have demonstrated that the expression of glutamine-rich PRD of human TIA-1 (hTIA-1) forms cytoplasmic microaggregates that sequester endogenous TIA-1 and prevents the formation of ARS-induced SGs [10]. In the present study, pctGFP-cTIA-1, pctGFP-cRRM, and pctGFP-cPRD plasmids were constructed and transfected to HeLa cells to evaluate the localization pattern of each domain. The results showed no SG dots in the pctGFP-cRRM-transfected cells. In ARS-treated cells, the overexpression of pctGFP-cRRM induced the formation of atypical SG dots that exhibited weak green fluorescence, which was localized with endogenous G3BP1 (panel A in Fig. 4; rows 3 and 6). In contrast, the overexpression of pctGFP-cPRD induced the formation of distinct cytoplasmic foci in the cells, even in the absence of treatment. Two types of cytoplasmic foci were observed in the pctGFP-cPRD-transfected cells. Approximately 55% of the pctGFP-cPRD-transfected cells harbored regular-sized cytoplasmic dots, whereas markedly larger dots were detected in the remaining ~45% of the transfected cells. More interestingly, the regular-sized dots, but not the large dots, colocalized with the endogenous SG marker (panel A in Fig. 4; rows 2 and 4).
Typical SGs disassemble in the presence of CHX, a drug that stabilizes polysomes and prevents protein translation [14]. Panel B in Fig. 4 shows that in ARS-treated cells, CHX induces the disassembly of SGs as indicated by the localization of endogenous G3BP1. As expected, the dots in the pctGFP-cTIA-1-transfected cells disassembled in the presence of CHX (panel B in Fig. 4; row 4). Similarly, in the ARS treatment group, the atypical SGs in pctGFP-cRRM-transfected cells also underwent disassembly upon CHX treatment. (panel B in Fig. 4; row 6). Furthermore, the large dots induced by pctGFP-cPRD transfection did not disassemble in the presence of CHX, whereas the regular-sized pctGFP-cPRD dots underwent disassembly (panel B in Fig. 4; rows 2 and 5). Collectively, these results indicate that pctGFP-cPRD induces the aggregation of large granules that differ from typical SGs.
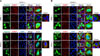
GFP tags influence the localization pattern of cPRD
Because the size of the GFP tag (238 amino acids [aa]) is much larger than that of HA (aa sequence: YPYDVPDYA) or the Flag tag (aa sequence: DYKDDDDK), this discrepancy may largely affect the localization of cPRD. To exclude artifactual localization due to the GFP tag, we constructed Flag-epitope-tagged cTIA-1, cRRM, and cPRD (Fig. 1). Fig. 4 shows that a similar localization pattern was observed in cells transfected with Flag-tagged cTIA-1/cRRM and those with transfected with pctGFP-tagged cTIA-1/cRRM. Next, we generated Flag-tagged and pntGFP-tagged cPRDs. Interestingly, no SG dots were observed in Flag-tagged cPRD-transfected HeLa cells in the absence of ARS (panel C in Fig. 5; row 3). On the other hand, transfection of pntGFP-tagged cells induced the formation of normal-sized SG dots (panel C in Fig. 5; row 2). When the cells were exposed to oxidative stress, typical SG dots were observed in both pFlag-cPRD- and pntGFP-cPRD-transfected cells (panel C in Fig. 5; rows 5 and 6). To examine whether the SG dots induced by cPRD transfection were canonical SGs, the transfected cells were treated with CHX. The results showed that only the large dots induced by pctGFP-cPRD failed to dissolve in the presence of CHX (panel D in Fig. 5; rows 4). In contrast, Flag-cPRD and ntGFP-cPRD induced the disassembly of dots upon exposure to CHX, indicating that the dots were typical SGs (panel D in Fig. 5; rows 5 and 6). Collectively, these results demonstrate that although the GFP tag did not influence the localization of cTIA-1 and cRRM, the C-terminal GFP tag has a notable effect on cPRD localization.
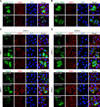
Endogenous cTIA-1 was aggregated as SGs in chicken cells and tissues under stress
To further elucidate the role of cTIA-1 in SG formation in chicken cells, endogenous TIA-1 was stained using an anti-TIA-1 antibody after exposure to various stressors. As expected, no SG dots were observed in the mock-treated DF-1 cells (panel A in Fig. 6; row 1). On the other hand, typical SG dots were detected in ARS-treated, heat-shock-treated, and virus-infected DF-1 cells (panel A in Fig. 6; rows 2–4, arrows). The expression levels of cTIA-1 mRNA in healthy chicken tissues were analyzed by performing qRT-PCR, and the results showed that cTIA-1 was constitutively expressed in all tissues examined, with highest levels observed in the intestine, kidney, and heart (panel B in Fig. 6). Next, we evaluated the formation of SGs in vivo under heat-shock stress. Panel C in Fig. 6 shows that in mock-treated chickens, cardiomyocytes were well arranged and contained no SG dots. In contrast, in heat-shock-treated chickens, cardiomyocytes were extensively damaged, and the perinuclear space had widened. As expected, distinct SG dots were detected in cardiomyocytes (panel C in Fig. 6; row 2, arrows). Collectively, these results demonstrate that endogenous cTIA-1 aggregates as SGs in vitro and in vivo.
Discussion
As an evolutionally conserved response to environmental stress, SGs have been detected in various species, including mammals, plants, and yeast [21018]. In human cells, TIA-1 is responsible for sequestering untranslated mRNAs at SGs in the cytoplasm. Therefore, TIA-1 is considered to have a key role in SG formation under environmental stress [15]. In chicken cells, TIA-1 has been shown to be essential for cell viability [16]. However, the role of TIA-1 during SG formation in chicken cells and tissues remains elusive.
The results of the present study have shown that under various stressors, the exogenous expression of GFP-tagged cTIA-1 leads to the formation of fluorescent dots in the cytoplasm of HeLa and DF-1 cells. Notably, the expression of cTIA-1 alone induced the formation of cytoplasmic TIA-1 dots in a small population of cells. Previous reports have shown that the expression of hTIA-1 resulted in the formation of SGs in 70% of transfected cells in the absence of stress [10]. Although transfection of cTIA-1 and hTIA-1 induced different amounts of SGs, the exogenous expression of cTIA-1 has resulted in the sequestration of endogenous TIA-1 and other SG markers as SGs.
Although some components of cytoplasmic granules vary depending on the type of stressor, the core components of SGs remain the same [121]. Typical SGs contain small ribosomal subunits, eIFs, and RNA-binding proteins. Moreover, canonical SGs dissolve upon treatment with CHX and emetine, which are drugs that stabilize mRNAs on polysomes [14]. Therefore, two experiments were performed to evaluate whether the cytoplasmic dots induced by pctGFP-cTIA-1 transfection were actual SGs. As expected, multiple SG markers such as G3BP1, PABP, and eIF3 were detected in cTIA-1 transfection-induced SGs. Moreover, pctGFP-TIA-1 dissolved in the presence of CHX, indicating that the overexpression of pctGFP-cTIA-1 induces the assembly of canonical SGs instead of merely forming cTIA-1 aggregates.
In most somatic cells, TIA-1 and TIAR accumulates in both the nucleus and the cytoplasm [15]. Moreover, these 2 RNA-binding proteins shuttle between these two subcellular regions. In this process, RRM2 and RRM3 function in nuclear accumulation and nuclear export, respectively [27]. The highest degree of identity among chicken, duck, human, and mouse TIA-1 is observed at the level of the RRMs (> 94%, data not shown). Therefore, the function of chicken RRMs is probably similar to that of human RRMs. As expected, GFP- or Flag-tagged cPRD, which lack RRMs, were mainly localized in the cytoplasm, particularly in cells under stress. Correspondingly, the transfection of GFP- or Flag-tagged RRMs resulted in a diffuse cytoplasmic and nuclear distribution. Moreover, similar to hTIA-1 RRMs, cRRMs did not affect the existing SGs under oxidative stress.
SG assembly is mediated by the PRD of TIA-1 in human cells [10]. In the present study, we first generated N-terminal-GFP-tagged cTIA-1 PRD and transfected this to HeLa cells. Both distinctly large- and regular-sized cytoplasmic dots were simultaneously detected in pctGFP-cPRD-transfected cells even in the absence of stress. The small dots were identified as typical SGs because they localized with endogenous G3BP1 and dissolved in the presence of CHX. In contrast, the large dots were devoid of endogenous G3BP1 and remained stable after exposure to CHX, indicating that these granules were not SGs. Previous reports did not observe such large granules in HA-tagged human-PRD-transfected cells [10]. Therefore, Flag-tagged and N-terminal GFP-tagged cPRD were constructed and transfected to cells. The results showed no large dots in the pctGFP-cPRD- and pFlag-cPRD-transfected cells, indicating that the C-terminal GFP tag did indeed affect cPRD localization. Several reports have shown that GFP-tagged proteins are functional and properly targeted [49]. In the present study, the GFP tag did not affect the localization of either TIA-1 or RRM, as shown by the same localization pattern between GFP- and Flag-tagged TIA-1s/RRMs, thereby indicating that both GFP-tagged TIA-1 and RRM were properly targeted. The size of the GFP tag (238 aa) was larger than the cPRD (95 aa), but smaller than cTIA-1 (373 aa) and cRRM (275 aa), which may partially explain the observed discrepancy in localizations. It is more likely that the property of the PRD is responsible for the formation of large dots. Proteins possessing PRDs have the ability to form aggregates. For example, in yeast, SUP35, which also possesses PRDs, can adopt two distinct conformations. SUP35 assumes a soluble conformation in the (psi−) strain, whereas it assumes an aggregation-prone conformation in the (psi+) strain [22]. Therefore, in the present study, the C-terminal GFP tag may be aggregated by cPRD to form large fluorescent dots. This process is independent of RNA translocation and protein translation because the dots did not dissolve in the presence of CHX.
It has been reported that endogenous chicken hsp24 is recruited in cytoplasmic aggregates in CEF cells [3]. In this study, we have shown that cTIA-1 is recruited as cytoplasmic aggregates under various stressors in chicken DF-1 fibroblast cells. Cells generally respond to heat-shock condition by expressing a series of heat-shock proteins [5]. Previous reports have shown that hsp27 and hsp70 protect the mammalian heart against myocardial infarction and ischemia-reperfusion injury, respectively [619]. Therefore, hearts of mock-treated and heat-shock-treated chicken were isolated to detect SG formation. Distinct SG dots were detected in cardiomyocytes of heat-shock-treated chicken indicating that cTIA-1 aggregates as SGs in vivo.
In conclusion, the findings of the present study demonstrate the role of cTIA-1 in SG formation in chicken cells and tissues; moreover, the cPRD has a role in SG aggregation. To evaluate the function of the PRD, the use of a C-terminal GFP tag should be avoided because it affects PRD localization.

 XML Download
XML Download