

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Over 500 O:H serotypes of Shiga toxin (Stx)-producing Escherichia coli (STEC) have been implicated in many human infections, including mild diarrhea, hemorrhagic colitis (HC) and hemolytic uremic syndrome (HUS) [41]. Although a single serotype, O157:H7 (referred to as O157 STEC), is most associated with outbreaks, epidemiological surveillance has reported that non-O157 STEC serotypes are responsible for approximately 30% of diseases in humans [14]. The consumption of raw or undercooked meats as well as water and other food products from fecal-contaminated environments with infected animals are common routes of transmission of STEC [11].
As a zoonotic bacterial pathogen, O157 STEC is characterized by (i) the production of one or more Stxs that have a cytopathic effect on mammalian cells, (ii) the presence of the locus of enterocyte effacement (LEE) that is responsible for attaching to and effacing lesions, and (iii) a 93 kilobyte virulence plasmid called pO157 [41]. Although a subset of non-O157 STEC strains have all the virulence factors typical of O157 STEC (previously defined as enterohemorrhagic E. coli [EHEC]), some strains are missing one or more such virulence factors except Stxs. Instead, they may possess additional adhesins and toxins, such as Saa (STEC auto-agglutinating adhesin [30]), Iha (IrgA homolog adhesin [36]), EcpA (E. coli common pilus [31]), CDT (cytolethal distending toxin [17]), and SubAB (subtilase cytotoxin [13]), to establish infection in the host's gastrointestinal tract. Large pO157-like virulence plasmids have also been reported among non-O157 STEC isolates [4].
Human infections by O157 or non-O157 STEC have been reported in many countries since the first documented O157 STEC fast-food-chain outbreak in 1982 in the USA [33]. In particular, 390 outbreaks have been caused by O157 STEC from 2003 to 2012, and 46 outbreaks were caused by non-O157 STEC from 1990 to 2012 in the USA [1523]. Notably, 93 and 104 outbreaks have occurred due to O157 and non-O157 STEC, respectively, in Japan from 2000 to 2012 [37]. In Spain, however, only 6 O157 and 2 non-O157 STEC outbreaks were reported from 1986 to 2007 [26]. Recently, the largest O104:H4 STEC outbreak occurred in Germany, and was responsible for 46 deaths, 782 cases of HUS, and 3,128 cases of acute gastroenteritis [12].
Cattle are the principal reservoirs for STEC, although STEC can colonize other animals (i.e., sheep, goats, pigs, horses, birds, fish, and rabbits) [21]. Thus, the prevalence of O157 or non-O157 STEC isolates in bovine feces and retail meats has been monitored annually in several countries, including the USA, Japan, Spain, and Argentina.
Although there have been a few asymptomatic or sporadic cases of STEC infection with mild diarrhea reported in Korea, no severe Korean STEC outbreaks in humans have been reported [22]. While the reasons for this are unclear, certain genetic variations, immune status, or dietary habitats unique to Korean people as well as distinct pathogenicities of endemic STEC isolates might be viable reasons. In our attempts to address this question, we isolated and characterized the virulence of seven STEC strains from 434 retail meats in Korea from 2006 to 2012. Phylogenetic relatedness was also examined by using multilocus sequence typing (MLST) and pulsed-field gel electrophoretic (PFGE) analyses.
Materials and Methods
Bacterial strains
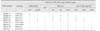
All E. coli strains in this study are listed in Table 1 [12038]. In general, bacteria were maintained in Luria-Bertani (LB; BD Diagnostics, USA) or Sorbitol MacConkey (SMAC; BD Diagnostics) media at 37℃. Seven STEC strains were isolated from 434 retail meat samples (beef, pork, and chicken) in Korea during 2006 to 2012. Each frozen stock sample was aseptically recovered on SMAC media at 37℃ for 19 h; single colonies were used for further experiments.
Sample collection and STEC isolation
STEC strains were isolated and identified according to the guidelines in the Standards for Processing and Ingredients of Specifications Livestock Products in Korea with slight modifications [27]. Briefly, 434 meat samples were collected from packaged raw beef, pork, and chicken meats from retail markets in Korea during 2006 to 2012. Twenty-five grams of each fresh meat sample was taken aseptically and homogenized with 225 mL of buffered peptone water (Oxoid, UK). For E. coli isolation, 10 mL of the homogenized mixture were blended in 90 mL of Brilliant Green Bile Broth (Oxoid) and incubated at 37℃ for 24 to 48 h. One loop of the enrichment culture was streaked onto Eosin Methylene Blue agar (Oxoid). Plates were examined for typical E. coli colonies after incubation at 37℃ for 18 to 24 h. From each positive sample, typical colonies were transferred onto Tryptone Soy Agar (TSA; BD Diagnostics) and incubated at 37℃ for 18 to 24 h. The resultant colonies were kept at −80℃ as 40% (v/v) glycerol stocks. Each isolate was identified by using the Vitek II Compact system (bioMé rieux, France). In addition, all isolates were confirmed by performing polymerase chain reaction (PCR) with primers specific to E. coli 16S rDNA and stx gene [27].
Determination of O and H types of STEC isolates
The E. coli O antigen types were determined by using a set of antisera against O1 to O185 according to the manufacturer's instruction (Laboratorio de Referencia de E. coli, Spain). For H antigen types, all H antigen-encoded fliC genes were amplified by using gene-specific primers, which are available upon request from the corresponding authors. After being cloned into a TA cloning vector (Real Biotech Corporation, Taiwan), the amplicons were sequenced and analyzed by using the BLASTN program (National Center for Biotechnology Information, USA).
Stx subtyping
To determine the Stx subtypes of each STEC isolate, the partial sequences of the stx1 and stx2 genes were determined and analyzed by using the BLAST program. The results were confirmed by using a multiplex PCR method as previously described [2].
Reversed passive latex agglutination assay
To quantitate Stx1 and Stx2 production in STEC isolates, we used a commercial reversed passive latex agglutination (RPLA) kit (VTEC-RPLA; Denka Seiken, Japan). A bacterial colony was inoculated into Tryptone Soy Broth (TSB; BD Diagnostics) and grown at 37℃ with vigorous shaking for 19 h. The bacterial culture was centrifuged for 20 min at 13,500 × g. Supernatants were obtained, and Stx titers were determined according to the manufacturer's instructions. Titers lower than 1:2 were interpreted as undetectable for Stx production.
Genetic prevalence of STEC-associated virulence factors
The presence or absence of other STEC virulence factors, previously defined among clinical STEC isolates, were determined in the isolates from this study. PCR was carried out by using target gene-specific primers, including cytotoxins (stx, subAB, and cdt [1317]), the LEE-encoded components for the type 3 secretion system (eae, tir, and espB [1928]), adhesins (saa, iha, and ecpA [2935]), and the pO157-encoded virulence factors (ehxA, espP, and katP [6828]). All primers used are available upon request from the corresponding authors. The resultant PCR products were visualized on a 1% agarose gel after ethidium bromide staining.
Acid resistance assay
Two important acid resistance (AR) phenotypes were evaluated in the STEC isolates from this study. One was the glucose-repressed RNA polymerase sigma factor (RpoS)-dependent AR phenotype (AR1), and the other was the glutamate-induced AR phenotype (AR2). In brief, a bacterial colony was inoculated into LB broth and grown at 37℃ with vigorous shaking for 19 h. The cells were diluted 1:300 in either HCl-acidified LB broth (pH 3.0; AR1) or minimal E medium containing 0.4% glucose media with 1.5 mM glutamate (pH 2.5; AR2) and further incubated at 37℃ with vigorous shaking for 2 h. The bacterial populations before and after the acid challenge were enumerated by direct plating. At least three independent experiments were performed, and the results are presented as mean ± SD.
Western blotting
Bacteria were cultured for 19 h at 37℃ with vigorous shaking as described above. The cells were collected by centrifugation and resuspended at 20 optical density at 600 nm (OD600) units per mL in 2× Laemmli loading buffer. After boiling for 10 min, the protein samples were separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were semi-dry transferred to polyvinylidene fluoride membranes (Immobilon-P; Bio-Rad, USA). Primary antibodies specific to RpoS (Santa Cruz, USA), GadA (glutamic acid deoxycarboxylase A; MyBioSource, USA), and DnaK (heat shock 70 kDa protein; Enzo, USA) were used, and anti-mouse rabbit IgG coupled to peroxidase (Enzo) was used as a secondary antibody according to the manufacturers' instructions. Blotted membranes were visualized by exposure to X-ray films by using the Western Detection Kit System (Abclon, Korea).
MLST analysis
The MLST analysis scheme uses internal fragments of the following seven house-keeping genes: adk, fumC, gyrB, icd, mdh, purA, and recA. These genes were amplified by PCR, and the products were analyzed by DNA sequencing. The sequences were submitted to the E. coli MLST database [40] to determine the corresponding sequence types (ST).
PFGE analysis
Standard PFGE analysis was carried out by using CHEF MAPPER (Bio-Rad). In brief, bacterial cells from an overnight culture in TSB were pelleted at 13,000 × g for 5 min. The pellet was embedded in 1.0% agarose plugs and lysed by proteinase K (Sigma-Aldrich, USA). Lysed plugs were then digested overnight with 40 U of XbaI (New England Biolabs, USA) at 37℃. Digested plugs were placed on 1.2% SeaKem Gold agarose (Lonza, Switzerland), and PFGE was carried out at 6.0 V for 19 h with an initial switch time value of 2.16 sec and final switch time of 54.17 sec in 0.5 × TBE (Tris/borate/ethylenediaminetetraacetic acid; Sigma-Aldrich) buffer at 14℃. PFGE patterns were analyzed by using Bionumerics software (ver. 6.5; Applied Maths, USA). The degree of similarity was calculated by using the unweighted pair group method with averaging based on Dice similarity co-efficiency with 2% of tolerance. More than three isolates with over 85% similarity were considered to be one cluster group.
Statistical analysis
Statistical analysis was performed via one-way ANOVA followed by a Tukey's multiple comparison test within OriginPro 8 software (ver. 8.0724; Origin-Lab, USA). For PFGE analysis, all statistical comparisons were performed by applying the Chi-squared test in SPSS software (ver. 12; SPSS, USA). P < 0.05 was considered statistically significant.
Results
Identification of STEC isolates
Among the 434 retail meat samples obtained during 2006 to 2012, 299 samples (68.9%) were culture-positive for E. coli and 7 samples (1.6%) were positive for STEC; of the seven STEC isolates, four were from beef samples and 3 from pork (Table 1). The STEC isolates belonged to four different serotypes: O91:H14 (3 isolates), O121:H10 (2 isolates), O91:H21 (1 isolate), and O18:H20 (1 isolate) (Table 2). The two O91:H14 beef-derived isolates had both stx1 and stx2b genes, whereas the O91:H14 pork-derived isolates had only the stx1 gene. The O91:H21 and O18:H20 beef isolates possessed the stx2c gene, while the two O121:H10 pork isolates possessed the stx2e gene (Table 2). The RPLA assay demonstrated that the individual STEC isolates produced different levels of Stx1 and Stx2. Compared to EDL933, the three STEC O91:H14 isolates secreted relatively high levels of Stx1, whereas most isolates except STEC O91:21 expressed undetectable levels of Stx2 (Table 2).
Presence of various virulence genes in STEC isolates
As determined via PCR analyses, none of the isolates carried the eae, tir, or espB genes, which are encoded in the LEE pathogenicity island. Instead, they all possessed one or more virulence genes for other alternative adhesins such as saa, iha, and ecpA (Table 3). Moreover, the two STEC O91:H14 beef isolates carried the subAB toxin gene. STEC O91:H21 and O18:H20 isolates carried the cdt toxin gene. STEC O91:H14 and O91:H21 isolates harbored one or more pO157-encoded virulence genes such as ehxA, espP, and katP (Table 3).
AR phenotypes of STEC isolates
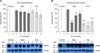
The survival of the STEC isolates under acidic conditions was evaluated based on their AR1 and AR2 phenotypes. As shown in panel A in Fig. 1, all STEC isolates could survive as well as EDL933 in the AR1 system (Table 1). No differences among the STEC isolates were observed in the expression of RpoS (panel A in Fig. 1; lower panel). However, we detected heterogeneity in survival rates among the STEC isolates in the AR2 system (panel B in Fig. 1; upper panel). A significantly reduced resistance was observed in five isolates under the AR2 system compared to that of EDL933 (p < 0.01). Interestingly, western blot analysis revealed that the observed AR2 phenotype did not agree with the expression profile of GadA (panel B in Fig. 1; lower panel).
Phylogenetic relatedness of STEC isolates
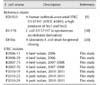
To assess phylogenetic relatedness among our STEC isolates, both MLST and PFGE analyses were carried out. As shown in Fig. 2, the MLST analysis revealed that the three STEC O91:H14 isolates were categorized into ST33, and the two STEC O121:H10 isolates were ST641. One O91:H21 beef isolate was ST442, while the O18:H20 isolate was un-typeable within the E. coli MLST database (see Materials and Methods section; Fig. 2). Subsequent PFGE analysis demonstrated that the two O91:H14 beef isolates shared an identical phylogenetic pattern, while the two O121:H10 pork isolates were closely related with a similarity of 88.2% (Fig. 2).
Identification of human STEC O91:H14 isolates in Korea
Since our current results showed the prevalence of 3 STEC O91:H14 among the 7 retail meat isolates of STEC (~42.9%), we hypothesized that this STEC serotype might be associated with human infections. To this end, we determined serotypes of 100 STEC isolates that were collected from human asymptomatic carriers (79 isolates) or diarrheal patients (21 isolates) in Korea during 2003 to 2014 (Table 4). Interestingly, the results showed that 96% (96 isolates) of them were identified as O91:H14, although a few other serotypes were present such as O141:H2, O141:Hnt, O146:H19, and O179:Hnt (Table 4).
We further investigated the genetic relatedness between our STEC O91:H14 retail meat isolates and the 18 human O91:H14 isolates from diarrheal patients by undertaking MLST analyses. As shown in Fig. 3, the results demonstrated that 13 of the 18 human STEC O91:H14 isolates (72.2%) from diarrheal patients belonged into the ST33 lineage, which was identical to that of the 3 retail meat isolates and the one human isolate from an asymptomatic carrier.
Discussion
In this study, we found a possible endemic STEC serotype in Korea, O91:H14. During the experimental period, STEC O91:H14 was isolated from retail meat samples; in addition, it was predominant in samples from human asymptomatic carriers and diarrheal patients. The retail meat isolates of STEC O91:H14 with ST33 poorly expressed Stxs, lacked the LEE pathogenicity island, and displayed a reduced AR2 phenotype in a GadA-independent manner, implying a low pathogenic potential. To our knowledge, this is the first report on an endemic STEC serotype in Korea, and the results imply a potential STEC link between retail meats and human infection.
A previous study suggested that STEC serogroup O91 strains should be seriously considered as human pathogens since they represent 35.7% of global STEC infections in humans [3]. Interestingly, only four O91 serotypes are associated with almost all STEC O91 cases (~90.7%), including STEC O91:H21 (39.2%), O91:H− (31.4%), O91:H14 (12.3%), and O91:H10 (7.8%) [3]. In this study, however, we detected STEC O91:H14 strains among the STEC isolates from the 464 retail meats (3 STEC O91:H14 among 7 STEC isolates) and the human patients (96 STEC O91:H14 among 100 STEC isolates) in Korea. Similarly, recent studies have reported that STEC O91:H14 strains are responsible for sporadic cases of human diseases in several countries [2425]. For example, Maeda et al. [24] showed that STEC O91:H14 is one of the major non-O157 STEC O serogroups commonly isolated from asymptomatic carriers in Japan. In Germany, STEC O91:H14 isolates are frequently associated with asymptomatic or mild diarrheal cases without HUS [25]. These results implied that STEC O91:H14 is rarely pathogenic in humans. Supporting this notion, 96.0% of the STEC O91:H14 isolates in Korea were recovered from asymptotic carriers or from sporadic cases of mild diarrhea. Although the reasons for such low pathogenicity are unknown, our beef and pork isolates of STEC O91:H14 (i) did not thrive in an acidic environment, despite encoding AR genes such as GadA from AR2 system [9], (ii) poorly expressed Stxs, and (iii) lacked the LEE pathogenicity island. Most importantly, all the STEC O91:H14 isolates from Japan [24], German [25], and Korea were categorized into the ST33 STEC lineage.
Several Stx variants have been reported based on the heterogeneity of their amino acid sequences, cytotoxic activities, and clinical manifestations [34]. Like EDL933, a human outbreak strain that has resulted in HC and HUS in the USA [1], our non-O157 STEC isolates had various combinations of Stx subtypes singly or doubly, but their expression levels of Stx subtypes were relatively lower than those of EDL933, especially in Stx2 variants (Stx2b, 2c, or 2e) being produced at undetectable levels. Although a single STEC O91:H21 isolate was identified as a high producer of Stx2c, as is EDL933, the isolate seems less tolerant to the acidic environments such as the AR2 system as well as less adherent to HeLA cells than EDL933 (Yoon et al., unpublished data). STEC O91:H21 harboring Stx2c has been isolated from sporadic HUS cases in humans [16]; thus, it would be interesting to see if our STEC O91:H21 isolate is pathogenic in vivo by using appropriate animal infection models.
In this study, the prevalence of non-O157 STEC from retail meats in Korea was approximately 1.6% (7/434 samples) during 2006 to 2012, which is equivalent to or slightly lower than previous studies in other countries. For example, the prevalence of STEC in retail beef samples has been reported to be 0.0%–1.1% for O157 and 0.0%–7.3% for non-O157 in the USA [51132], 1.0% for O157 and 11.0% for non-O157 in Spain [32], and 12.2% for O157 and 14.4% for non-O157 in Argentina [7]. Similar to results from beef, a recent study suggested that pigs were important reservoirs of STEC by demonstrating that approximately 4.59% of STEC strains were isolated from 764 samples across the pork production chain in Argentina [10].
DNA fingerprinting analysis provides useful information on the phylogenetic relatedness among bacterial strains. In this study, all three STEC O91:H14 isolates were identified as single-locus variants belonging to ST33; similar to that reported for STEC O91:H14 isolates from asymptomatic carriers or non-HUS patients in Japan or Germany [2425]. Moreover, two of these isolates shared identical molecular patterns as per our PFGE results, implying that both strains are a possible clonal set with the same evolutionary origin. A recent study showed that STEC O91 serogroups with ST442 are associated with HUS patients [25]. Therefore, we speculated that different lineages of STEC O91 isolates might possess distinct pathogenicity or host adaptive capabilities in humans or animals.
Previous studies have demonstrated that STEC serogroup O91 isolates are prevalent among food sources [1839] as well as in human patients [1839] in other countries. This type of prevalence is also likely in Korea because our data indicated a high prevalence of STEC O91:H14 in human carriers and/or patients (96.0%). It should be noted that STEC O91:H14 isolates with the ST33 lineage were prevalent in both retail meats and human diarrheal patients in Korea although only limited information is currently available.
In conclusion, this is the first report showing that STEC O91:H14 belonging to ST33 is prevalent among retail meats and human diarrheal patients in Korea. Based on our data, we propose STEC O91:H14 with ST33 as an endemic non-O157 STEC clone with a likely STEC link between retail meats and human infection in Korea. Further study is warranted to confirm that STEC O91:H14 with ST33 is a dominant serovar among humans, retail meats, and animal feces in Korea as well as to determine their virulence potentials and host adaptability in vivo.




 XML Download
XML Download