

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Degenerative mitral valve disease (DMVD) is defined as the progressive degeneration of the mitral valve [4]. Mitral valve leaflets, which are naturally thin, translucent, and soft, become thickened and elongated with disease progression, leading to mitral regurgitation [7]. DMVD is the most commonly acquired cardiac disease in dogs [7]. The disease is usually found in small breeds; large breeds are apparently less prone to DMVD [8]. The disease is age-dependent, with increasing prevalence in dogs that are 7 years of age and older [4]. The etiology of canine DMVD remains unclear [3]. Recent evidence from highly susceptible breeds, including Cavalier King Charles spaniels and dachshunds, shows a strong inheritance pattern for DMVD with polygenic inheritance [2327] The breed specificity of DMVD incidence could be considered as evidence of a genetic basis for this disease in dogs because modern domestic dogs could easily sustain a detrimental genetic mutation [26].
Serotonin has a crucial role in various cardiovascular disorders. The carcinoid syndrome, which is developed by the secretion of serotonin and other vasoactive substances from a tumor, results in endocardial damage [21]. Furthermore, components of the cardiovascular system, such as vascular endothelium, smooth muscles, and heart tissue, easily bind to and transport serotonin. This increases serotonin signaling or decreases serotonin clearance, which may induce valvular lesions [1125]. In a previous study, rats that received serotonin injections over a long period developed valvular lesions [14]. Cardiac fibrosis and valvulopathy occur in mice without serotonin transporter (SERT) protein expression through an increase in serotonin production, localized serotonin re-uptake by SERT, and a decrease in serotonin metabolism [20]. Furthermore, carcinoid tumors that produce serotonin and the administration of serotonergic drugs have been shown to be associated with valvulopathy in humans [28]. In addition, expression of the SERT gene (SLC6A4) was significantly higher among dogs with clinical DMVD [10]. SERT expression facilitates the intercellular processing of serotonin after receptor interactions [221] and is responsible for serotonin uptake and consequent inactivation of amine. A previous study showed that interference with serotonin transmembrane processing via knocking out SERT gene resulted in valvulopathy in mice, establishing a link between SERT and the development of cardiac fibrosis and valvulopathy in vivo [621]. Absence of transmembrane processing may result in increased and persistent serotonin receptor interactions and increased valvular mitogenic activity and extracellular matrix production in SERT-knockout mice [1421].
Several studies have reported the molecular mechanisms underlying DMVD pathophysiology [24]. In human and veterinary medicine, serotonin and serotonin-related pathways have been associated with cardiac disease [1620]; serotonininduced valvular lesions resemble many gross and histologic DMVD descriptions, and serotonin has recently been suggested to have a role in the progression of DMVD in dogs [1132]. Histopathologic changes in the valves of DMVD dogs include damage to the endothelial valvular cell lining [9]. Furthermore, a phenotype transformation occurs in fibroblast-like valvular interstitial cells, which are responsible for normal valve structure via maintenance of the extracellular valve matrix, that change into a more active myofibroblast phenotype [6]. Serotonin has been suggested to both directly and indirectly stimulate such a phenotype transformation [2]. According to a previous study, the activated myofibroblasts can induce signaling pathways, resulting in the degenerative structure of the valve [6].
In this study, we searched for genetic variations in the SERT gene in Maltese dogs with DMVD. In addition, we investigated the possibility of changes in protein functions to reveal potential pathogenicity. Taken together, the relationship between canine DMVD and genetic variation was evaluated.
Materials and Methods
Animals
This study was approved by the University of Konkuk Institutional Animal Care and Use Committee (IACUC No. KU15115). Informed owner consent was obtained. All dogs were selected from those referred to the Veterinary Medical Teaching Hospital of Konkuk University. All 30 dogs in this study underwent echocardiographic examination with measurement of the cardiac function criteria. The right parasternal 4-chamber view was used for evaluation of mitral valve degeneration and detection of mitral regurgitation via color Doppler sonography. Based on the examination results, 20 dogs were diagnosed with DMVD and enrolled in this study. Additionally, 10 Maltese (all < 10 years of age) without mitral valve degeneration were included as control dogs. Maltese dogs were excluded if they had a congenital heart disease, other acquired cardiovascular disorders, or systemic organ-related diseases.
The 20 dogs diagnosed with DMVD were classified into groups based on heart disease stages B, C, and D of the American College of Veterinary Internal Medicine (ACVIM) classification [3]. The stage B group included Maltese dogs with DMVD that had never developed clinical signs caused by heart failure and they have only typical murmur; the stage C group included those with past or current clinical signs of heart failure, including exercise intolerance, coughing, labored respiration and dyspnea; and the stage D group included those with end-stage disease and clinical signs of heart failure that were refractory to standard treatment including furosemide, angiotensin-converting enzyme inhibitor, and pimobendan. Maltese dogs in stage A were excluded because that group includes dogs at high risk of developing heart disease but lacking an identifiable structural disorder such as a heart murmur.
Maltese dogs with DMVD, but suspected to have a systemic disease, including neurological, adrenal, thyroidal, renal, or hepatic disease and neoplasia, were excluded. Dogs suspected of having DMVD but did not fulfill all aspects of the inclusion criteria were excluded. Ten healthy Maltese dogs without pathologic findings evident in the same diagnostic examinations were included as the control group.
Blood sample collection and processing
Blood was collected from the jugular vein into 1.5 mL ethylenediaminetetraacetic acid tubes. Genomic DNA was extracted from whole-blood leukocytes by using a DNA Blood Mini Kit (Qiagen, Germany) and stored at −20℃. The DNA quality and purity were evaluated by using a NanoDrop 2000 spectrometer (Thermo Scientific, USA). All genomic DNA samples were confirmed by using gel-based polymerase chain reaction (PCR) with GAPDH primers (5′-GGT CAC CAG GGC TGC TTT-3′ and 5′-ATT TGA TGT TGG CGG GAT-3′). PCR for SERT was performed with primers used previously [32]. Table 1 shows the details of the primer sequences. PCR reagents were prepared as following conditions: 50 µL PCR volume with 10 µL of buffer containing 300 mM Tris-HCl (pH 9.3); 300 mM salts containing K+ and NH4+, 20 mM Mg2+, 5 mM dNTP, 20 pmol of both primers, and 1 µL of genomic DNA. The thermocycler profile was as follows: 5 min at 95℃, followed by 40 cycles of 30 sec at 95℃, 30 sec at 60℃, and 30 sec at 72℃ and a final extension step of 3 min at 72℃.
Sequencing and genotyping
Single nucleotide polymorphism (SNP) genotyping in the DMVD dogs, performed by DNA sequencing of PCR products that were generated with same primer pairs, was performed in a commercial laboratory (Cosmogene Tech Laboratories, Korea). The next step included a BLAST search for SLC6A4 (SERT gene) sequences in dog-specific databases at the National Center for Biotechnology Information (NCBI) website by using mRNA sequences with accession numbers NM_001110771.1 (Canis lupus familiaris solute carrier family 6 [neurotransmitter transporter, serotonin], member 4 [SLC6A4]) and whole-genome shotgun DNA sequences with NC_006591.3 (Canis lupus familiaris breed boxer chromosome 9, CanFam3.1, wholegenome shotgun sequence) as queries. These sequences were aligned for searching variations between cases and standard sequences. CLC sequence viewer (ver. 7; Qiagen) was used for the alignment of multiple sequences.
Structural modeling and polymorphism prediction
DNA sequences were imported and aligned by using CLC sequence viewer (ver. 7). Exonic regions were evaluated to identify the SNPs. Protein structure modeling and structure predictions of the polymorphisms were evaluated by using ModWeb (ver. r189; University of California at San Francisco, USA) and RaptorX [18]. Finally, the functional effects of the polymorphisms were predicted by using Polymorphism Phenotyping ver. 2 (PolyPhen-2) [1].
PolyPhen-2 calculates the possibility that a given mutation is damaging and shows estimates of false-positive (the chance that the mutation is classified as damaging when it is not) and true-positive (the chance that the mutation is classified as damaging when it is indeed damaging) rates. A mutation may also be qualitatively appraised as benign, possibly damaging, or probably damaging [1]. A previous study of canine genetic disease used PolyPhen-2 to predict non-conservative amino acid exchange effects on protein function [13].
ModWeb is a server providing for automated comparative protein structure modeling. It accepts sequences in the FASTA format and calculates models based on the best available template structures. Sequence-structure matches are established by using multiple variations of sequence-sequence, profilesequence, sequence-profile, and profile-profile alignment methods. Significant alignments (E-values > 1) covering at least 30 amino acid residues are selected for modeling. The models are built for each sequence-structure match by using comparative modeling with satisfaction of spatial restraints as implemented in Modeller [29]. The resulting models are evaluated using several model assessments [33].
RaptorX, a protein structure prediction server, predicts three-dimensional (3D) structures of protein sequences. For a given FASTA sequence, RaptorX predicts the secondary and tertiary structures as well as the accessibility and disordered domains. RaptorX also calculates p values for relative global quality, global distance test (GDT), and unnormalized GDT (uGDT) for absolute global quality in the model. The 3D structures can be visualized through a prediction server [5].
Results
Group characteristics
The mean age of DMVD group was 10.7 ± 0.6 years (each ACVIM stage: stage B, 11.1 ± 1.5 years; stage C, 10.3 ± 1.4 years; and stage D, 10.7 ± 2.5 years), while that of the control group was 11.9 ± 2.88 years. Male dogs comprised 53.1% of the DMVD group and 60.0% of the control group. According to the ACVIM staging system, seven dogs were stage B, ten were stage C, and three were stage D (Table 2).
Sequencing and genotyping
DNA sequence analysis of 13 SERT gene exons from the 20 DMVD-affected Maltese dogs and 10 control dogs was performed. DNA extraction was confirmed by observing positive PCR amplification of GAPDH, a housekeeping gene. All 13 exons of the canine SERT gene from all dogs were amplified by using each primer pair. Sequences from the groups were aligned with the reference canine SERT sequence by using the CLC sequence viewer. All sequences were confirmed by undertaking chromatogram analysis. The sequences of the control group revealed that the SERT gene sequences of the control group were identical with that of the reference canine SLC6A4 sequence (NM_001110771.1) from the NCBI database. When SERT sequences from the DMVD group compared with the reference sequence, genetic variations were detected in five exons (3, 4, 5, 7, and 9), one of which was synonymous and five nonsynonymous (Table 3).
Polymorphism prediction and structural modeling
Five nonsynonymous SNPs were identified in 10 of the Maltese dogs with DMVD. Of the five, two and three were predicted to be “benign” and “probably damaging,” respectively, to protein structure and function, based on the results of analysis using software to predict damaging missense mutations (Table 3). The samples from seven Maltese dogs contained SNPs considered “probably damaging.” The mean age of the seven dogs with SNPs predicted as probably damaging was 11.5 ± 1.2 years, while that of the 13 DMVD dogs with no mutation or a mutation predicted as benign was 10.1 ± 1.6 years; a statistically significant difference (p = 0.04).
Analysis using the ModWeb server confirmed the protein prediction model results were reliable with the following threshold scores: ModPipe quality score > 1.1, GA341 model score > 0.7, E-value < 0.0001, and z-discrete optimized protein energy < 0. All prediction model results were within the reliable ranges in ModWeb.
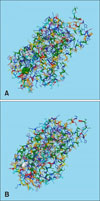
The 3D structure of the native model was generated by using RaptorX (Fig. 1). The SERT gene contains 630 amino acid residues. Of these, 599 amino acid residues (88%) were modeled, and 81 to 83 amino acid residues (12%–13%) of the whole SERT gene were predicted to be disordered. Analysis of the predicted secondary structures revealed 45% to 62% alpha helices, 4% to 11% beta sheets, and 32% to 42% loop structures. In particular, the ratio of the most stable alpha helix structure was decreased in the thymine deletion mutation. The results imply that mutation affects the whole structural composition of the SERT gene. The overall uGDT value range was 379 to 458 (61%–72%). The uGDT measures absolute model quality, and an uGDT > 50 is a reliable indicator of protein modeling. The p value reflects the likelihood of a predicted model being worse than the best of a set of randomly generated models for this protein sequence; therefore, p values indicate the relative quality of a model. The smaller this value, the higher the quality of the model. For alpha proteins, p values less than 10−3 indicate good quality. Similarly, p values less than 10−4 in mainly beta proteins indicate good quality. In this study, the RaptorX-predicted p values were 1.35e−16 to 5.87e−15. Five mutant models were generated using RaptorX (Table 4).
Discussion
Recent studies on the genetic aspects of DMVD have used various approaches [78]. The results of previous studies suggest that elevated serotonin signaling or decreased SERT function can activate signaling pathways in canine mitral valve disease. Another study reported decreased manifestation of the serotonin uptake transporter in dogs with DMVD when compared to unaffected dogs [12]. Previous studies revealed that the expression and activity of the SERT protein in humans are influenced by genetic variations in the SERT gene (SLC6A4) and that these genetic variations may influence susceptibility to adverse phenotypes associated with serotonin signaling [1731]. In particular, a 44 bp insertion or deletion in the SERT promoter region regulates transcriptional efficiency, with the short variant demonstrating 40% to 70% reduction in SERT expression and a 30% to 40% decrease in SERT protein levels [19]. Platelets containing rich SERT proteins have demonstrated differential rates of serotonin uptake, depending on the presence of SERT gene variations in dogs [15]. On the basis of these studies, we searched for variations in the SERT gene in Maltese dogs with DMVD. To our knowledge, this is the first study genotyping the SERT gene in a specific canine species via clinical sampling.
In this study, sequencing of the SERT gene in 20 dogs with DMVD confirmed the presence of variations in the gene. We identified six SNPs in five exons in the DMVD group. The results suggest a linkage between SERT polymorphism and DMVD. Additional analysis with prediction programs revealed that three of the six SNPs were “probably damaging” to the predicted protein function. In particular, 1193delT (thymine deletion at position 1193) was identified in seven Maltese dogs with DMVD, and a potential association of that SNT with the disease was suspected but was not statistically verified.
Previous study showed that decreased SERT expression occurs in valve tissue during late-stage canine DMVD [30]. Downregulation of SERT could have an important exacerbating role in the pathogenesis of late-stage disease, but the specific mechanism underlying SERT downregulation in DMVD is unknown. Decreased SERT expression is suspected to affect serotonin clearance. However, we could not directly confirm serotonin receptor protein expression; therefore, several prediction methods were used to validate the SERT gene variations.
Secondary and tertiary structure models of the SERT variants were identified in this study. Those models could serve as starting points for further analysis in a number of diverse applications. For example, the predicted 3D models could be used to predict substance-binding sites and epitopes. Additional applications include determining the binding of small ligand molecules to domain-binding sites [34]. Such moleculardocking studies could be performed by using software and, often, can have a critical guiding role in rational drug design [30]. A related principle, macromolecular docking, refers to the computational modeling of the quaternary structure formed by two or more protein domains [22]. Our study provides basic information for further protein-protein interaction studies.
In conclusion, this study identified six polymorphisms of the SERT gene in Maltese dogs with DMVD. Three of the six polymorphisms were predicted to be probable causes of damage to protein function. Confirmation of these predictive models, including the secondary and tertiary structures, was assessed by using various protein modeling standards. In addition, this study identified a 1193delT polymorphism in the SERT gene in Maltese dogs with DMVD, and that polymorphism was revealed as evidence of an association between the SERT gene and canine DMVD. Therefore, genetic analysis of the SERT gene in Maltese dogs is suggested for acute diagnosis of DMVD.




 XML Download
XML Download