

 PDF
PDF Citation
Citation Print
Print


Verminous encephalomyelitis is a neurologic disease in horses caused by the migration of nematodes and fly larvae. Equine cerebrospinal nematodiasis is associated with Parelaphostrongylus (P.) tenuis, Setaria digitata, Strongylus vulgaris, Strongylus equinus, Angiostrongylus cantonensis, Halicephalobus gingivalis, and Draschia megastoma [15]. Larvae of Hypoderma spp. have also been identified as a cause of equine verminous encephalomyelitis [5]. The meningeal worm, P. tenuis, commonly affects white-tailed deer and, rarely, sheep, goats, cattle, antelope, black-tailed deer, moose, elk, reindeer, fallow deer, and llamas [5]. Recently, verminous encephalitis caused by Protostrongylidae was identified in a horse based on the morphological characteristics of the nematodes and their distribution in the central nervous system (CNS) [7]. Equine meningeal worm infections have previously only been documented in the USA [7,8]. In the current study, we describe equine eosinophilic encephalomyelitis produced by nematodes, presumably P. tenuis.
Four thoroughbred horses presented neurological signs for two months from the end of August to October. The affected animals were kept at a private riding club in central (three animals) or southern (one animal) Korea. In August, the owners reported clinical signs, including postural abnormalities. Although the animals had normal appetite, they became thin due to dysbasia (Table 1). No other clinical signs were observed. Additionally, the four horses had been raised only in Korea and had not moved to or from other countries. Eventually, the animals were humanely euthanized, due to poor prognosis, and necropsies were performed.
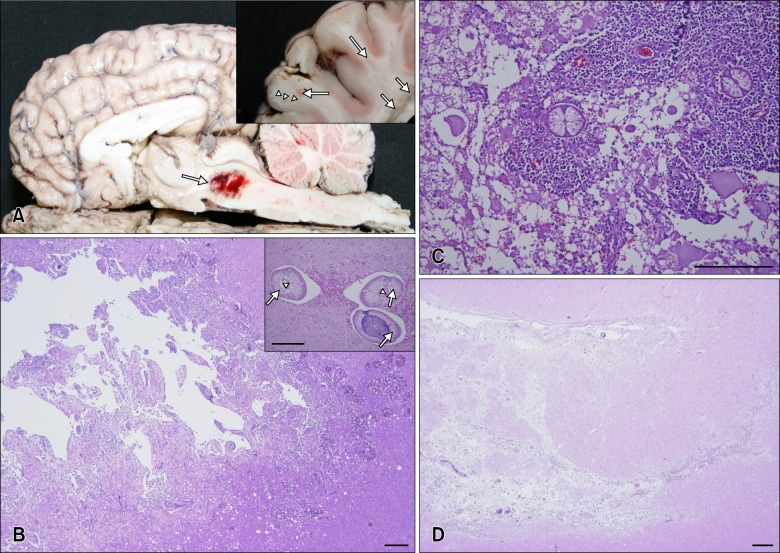
The cerebrum, midbrain, and spinal cord had gross, pale- to dark-red manifestations (panel A in Fig. 1). No other gross lesions were observed. Samples from brain, spinal cord, lungs, heart, liver, spleen, kidneys, lymph nodes, muscles, and intestine were collected, fixed in 10% buffered formalin, embedded in paraffin, and sectioned at 4 µm. The sections were then stained with hematoxylin and eosin (H&E) and Giemsa solution for histopathologic and parasitologic examination, respectively. Additionally, fecal samples were collected for fecal testing, and brain samples were cultured on both blood and MacConkey agars for bacterial isolation. No parasitic eggs or bacteria were detected.
Lesions were only observed histopathologically in the CNS. Multiple migration tracts existed in the white matter, the junction between the grey matter and white matter of the cerebrum, the brain stem, and, occasionally, extended into the grey matter. There was abundant eosinophil infiltration around blood vessels, as well as a few lymphocytes and macrophages. Hemorrhage, vacuolation, axonal spheroids, and intact or degenerated nematodes were observed in and around the migration tracts of the brain and/or spinal cord (panels B and C in Fig. 1). Multinucleated giant cells, macrophages, lymphocytes, and eosinophils were also observed around the degenerated nematodes. The nematodes had polymyarian-coelomyarian musculature, a smooth thin cuticle, and intestines lined by multinucleated cells with microvilli (panel B in Fig. 1). Compared to the brain, less severe histological features were present in the spinal cord. Larvae were observed in the thoracic spinal cords (panel C in Fig. 1), and migration tracts were present in the lumbar spinal cord. Additionally, hemosiderincontaining macrophages were observed in chronic lesions along with a connective tissue increase (panel D in Fig. 1).
Based on the observed morphological characteristics of the nematodes in the affected horses, the adult worm and larvae were classified to the superfamily Metastrongyloidea [2]. Within this group, parasite families that are known to invade the CNS include the Protostrongylidae, Angiostrongylidae, and Elaphostrongylinae [1]. Among those families, species in the Protostrongylidae and Angiostrongylidae families are reported to infest horses [17]. Some nematodes are host-specific, such as Angiostrongylus (A.) cantonensis (Angiostrongylidae), which, in rats, infests the CNS and develops from third-stage larvae to adult parasite. However, in aberrant hosts, migrating A. cantonensis larvae fail to mature in the CNS and remain in the larval stage [1]. Histopathological lesions in the CNS of aberrant hosts are less acute, due to mechanical stimuli, than severe granulomatous encephalomyelitis, probably due to responses to excretory materials and molting [9]. Additionally, Elaphostrongylus spp., such as Elaphostrongylus (E.) rangiferi and E. cervi, exhibit neurotropism and can cause neurological problems in aberrant hosts, such as sheep or goats, or patent hosts, such as reindeer and red deer. During aberrant host infestation, there are fewer adult Elaphostrongylus worms than infective third-stage larvae (L3) present in the CNS [34]. In this study, adult parasites were observed in cerebrum of all four horses but larvae were only observed in spinal cord.
Three species of the genus Parelaphostrongylus, including P. andersoni, P. odocoilei, and P. tenuis, are known to infect ungulates naturally, including bovids and cervids [1]. P. andersoni infects muscles of the hindbody including the longissimus dorsi, gluteal, thigh, and psoas, as well as lung, fat, and lymph nodes in white-tailed deer and caribou in North America and Eurasia [1]. P. odocoilei infection is typically observed in the skeletal muscles of black-tailed deer and mule deer in North America. P. tenuis is a common neurotropic nematode parasite of white-tailed deer in eastern North America [1]. However, in Korea, there are no known natural hosts of the genus Parelaphostrongylus until the equine observations reported herein. In Korea, roe deer (Capreolus pygargus tianschanicus) and water deer (Hydropotes inermis argyropus) are widely distributed in the wild. Given the presumed equine Parelaphostrongylus infections, a survey of Parelaphostrongylus infection in roe and water deer is required to determine its route of infection.
Adult P. tenuis are typically observed in the cranial subarachnoid space and dural venous sinuses. Mature female worms deposit eggs into the venous circulation, occasionally on the meninges or in the venous sinuses [1]. These eggs can then be carried hematogenously to the lungs, where they develop to first-stage larvae (L1) that can avoid granulomatous inflammation. The L1 travel up the host airway and exit via host feces. Free-living L1 infect terrestrial gastropods and develop into L3 in intermediate hosts [1]. Definitive or aberrant hosts become infected through accidental ingestion of intermediate hosts with L3. The L3 then migrate from abomasum to the spinal cord and are carried through the parenchyma of the spinal cord to the brain [1]. In this study, L3 were observed in the lumbar spinal cord of the infected horses.
It has been suggested that young calves are more susceptible to Parelaphostrongylus infection than adult cattle [6]. However, animal susceptibility to an aberrant infection by Parelaphostrongylus spp. could be different among the species. In the current study, Parelaphostrongylus infection was observed in adult horses.
Based on the morphological characteristics of the parasites and the infection sites, the present study presumes P. tenuis to be the causative agent producing CNS problems with ataxia and paresis in four horses, though a specific identification was not determined. Given that this is first documented case of P. infection in Korea, the authors suggest that Parelaphostrongylus infection should be added as a differential diagnosis for neurological problems in horses in Korea.
 XML Download
XML Download