

 PDF
PDF Citation
Citation Print
Print


Introduction
Amyotrophic lateral sclerosis (ALS) is a neurodegenerative disease associated with the selective motor neuron death in the spinal cord, brain stem, and motor cortex. Typical ALS patients suffer from a gradual loss of motor function caused by muscle atrophy and degeneration. To date, the main cause of ALS has not been identified, but various pathological hallmarks such as misfolded protein aggregation, mitochondrial dysfunction, glutamate excitotoxicity, and neuro-inflammation seem to be related to the development of ALS symptoms [82831]. Most cases of ALS are classified as sporadic ALS (sALS), while 10% of ALS cases are inherited familial ALS (fALS). To date, studies of fALS have identified many causes of the disease such as mutations of superoxide dismutase 1 (SOD1) [32], TAR DNA binding protein (TDP-43) [29], and translocated in liposarcoma/fused in sarcoma (FUS/TLS) [21]. The SOD1, a principal intracellular antioxidant enzyme, converts superoxide radicals to hydrogen peroxide, and SOD1 mutation can lead to multiple pathogenic mechanisms in ALS especially oxidative stress [5]. In previous study, many analyses of oxidative stress in ALS have been carried out to elucidate the relationship between degenerated motor neurons and elevated oxidative stress in ALS, and the results provide evidence supporting indications that oxidative stress affects motor neuron death and other cells [56]. In particular, microglia have a key role during the development of neurodegeneration by catalyzing the conversion of superoxide anion to hydrogen peroxide and inducing oxidative stress in the central nervous system (CNS) of ALS [4]. In particular, under certain pathological conditions microglia produce a high level of reactive oxygen species (ROS) through gp91phox, which is one of the main sources of ROS in microglia [132440]. The increase in ROS by gp91phox amplifies signals to produce neurotoxic factors through the MAPK pathway. Consistently, deletion of gp91phox, which is overexpressed in ALS microglia, can improve survival of hSOD1G93A Tg mice [172636].
Until now, various therapeutic approaches including stem cell-based clinical trials and adjustment of drug treatments have been attempted, but no significant effective and feasible therapies have been developed [83038]. The novel chemical compound KCHO-1 is a natural ethanol extract obtained from traditionally used herbal combinations which include Curcuma (C.) longa, Salvia (S.) miltiorrhiza, Gastrodia (G.) elata, Chaenomeles sinensis, Polygala (P.) tenuifolia, Paeonia japonica, Glycyrrhiza uralensis, Atractylodes (A.) japonica and processed Aconitum carmichaeli, and this combination of nine herbs has been used in traditional medicine [1423].
In this study, we aimed to determine whether KCHO-1 can reduce oxidative stress and prevent degeneration of motor neurons in an ALS model. To address this issue, we orally administrated KCHO-1 to hSOD1G93A Tg mice. The KCHO-1 administration delayed disease onset and improved motor activity in symptomatic hSOD1G93A Tg mice. Importantly, we found that KCHO-1 could reduce oxidative stress by reducing gp91phox expression and affecting the MAPK pathway in spinal cords of hSOD1G93A Tg mice, suggesting the therapeutic potential of KCHO-1 for ALS treatment.
Materials and Methods
Animals
The hSOD1G93A Tg mice are hemizygous transgenic B6SJL mice that carry a mutant human SOD1 gene, which has a glycine to alanine base pair mutation at the 93rd codon of the cytosolic Cu/Zn SOD1 [B6SJL−Tg(SOD1 × G93A)1Gur/J]. The transgenic mice used in this study were purchased from the Jackson Laboratory (USA), and that lab specified that the transgenic mice used in this study have an average lifespan of 128.9 ± 9.1 days. For selection of hSOD1G93A Tg mice by genotyping, DNA was extracted from the tail of the mice, and a PCR assay was performed. All mice were housed at a constant temperature of 21℃ to 23℃ and humidity of 50% to 60% under a 12 h light/dark cycle. Female hSOD1G93A Tg mice were colonized per experimental group (n = 11–12). Mice generally developed disease signs of neuromuscular deficits at an age of 105 to 115 days. All animal experiments were consistent with the guidelines of the Institute of Laboratory Animals Resources (SNU-120821-5-1; Seoul National University, Korea).
Materials
The KCHO-1 used in this study was consistent with that described in a previous study [23]. Briefly, The KCHO-1 was a mixture of herbs, which consists of C. longa (HP2013-10-01), S. miltiorrhiza (HP2013-10-02), G. elata (HP2013-10-03), Chaenomeles sinensis (HP2013-10-04), P. tenuifolia (HP2013-10-05), Paeonia japonica (HP2013-10-06), Glycyrrhiza uralensis (HP2013-10-07), A. japonica (HP2013-10-08), and processed Aconitum carmichaeli (HP2013-10-09). The C. longa (4 kg), S. miltiorrhiza (4 kg), G. elata (4 kg), Chaenomeles sinensis (2 kg), P. tenuifolia (2 kg), Paeonia japonica (2 kg), Glycyrrhiza uralensis (2 kg), A. japonica (2 kg), and processed Aconitum carmichaeli (1 kg) were mixed and extracted in 30% ethanol for 3 h at 84℃ to 90℃. The extract was concentrated and lyophilized by using a rotary evaporator. Four marker compounds were selected and selection criteria for the compounds were based on the quantitative method of the Korean Pharmacopeia (Table 1). All specimens were processed and deposited at Hanpoong Pharm & Foods (Korea) [1423].
Oral delivery of KCHO-1
Toxicity testing of oral-delivered KCHO-1 was previously studied, and there were no reported side effects at the high dose level of 2,000 mg/kg [14]. In this study, KCHO-1 at a dose of 250 mg/kg was treated daily. The age of the transgenic mice at the start of KCHO-1 treatment was 13 weeks and KCHO-1 was administered daily for 4 weeks (mouse ages from 91 days to 119 days).
Behavioral analysis
Before KCHO-1 was delivered to mice, behavioral tests were performed. Prior to testing, the mice underwent a one-week training period to adapt to the apparatus (7650 Accelerating Model, Ugo Basile, Italy). After the training period, 13-week-old mice were subjected to a rotarod test. The motor function of a mouse was assessed by measuring the time that a mouse could remain on the apparatus at a rotation rate of 10 rpm. Tests were performed every other day, and the average time for three attempts was recorded for each mouse.
Cell viability assay
The BV2 microglial cells were maintained in Dulbecco's modified Eagle's medium (11995; Gibco, USA) supplemented with 10% FBS (16000; Gibco), 1% penicillin and streptomycin (15140; Gibco). The cells were seeded in 24-well plates. After stabilizing, several concentrations of KCHO-1 were treated (3.1–100 µg/mL). After 24 h, 5 mg/mL MTT solution (Sigma, USA) was treated and the cells were incubated for 4 h at 37℃. The supernatant was discarded and DMSO was added to the BV2 microglial cells to solubilize the formazan crystals. Subsequently, the supernatant was moved to 96-well plates and absorbance was measured by using an EL800 microplate reader (BioTek Instruments, USA) at a wavelength of 540 nm.
Griess assay and ROS test
The BV2 cells were seeded in 24-well plates. After stabilization, the cells were treated with several concentrations (3.1–100 µg/mL) of KCHO-1, and the cells were activated simultaneously with 10 ng/mL lipopolysaccharide (LPS; Sigma) and 10 ng/mL interferon (IFN)-γ (PeproTech, USA). After 24 h, the supernatant was used to determine the nitrite concentration, which was measured by using a colorimetric Griess reagent system according to the manufacturer's instructions (TB229; Promega, USA). Attached cells were used in a ROS test, in which, the cells were treated with CellROX oxidative stress reagents (C10422; Life Technologies, USA) for 30 min at 37℃ and then detached by pipetting. Harvested cells were analyzed by performing flow cytometry (BD Bioscience, USA). The ROS test of the spinal cord of hSOD1G93A Tg mice was performed by using H2DCFDA. Cryotissues were sectioned at 10 µm and then stained with H2DCFDA (D399; Thermo Fisher, USA) by treating with 5 µM H2DCFDA for 30 min at 37℃. After 30 min, the ROS level were measured by using an Eclipse TE200 confocal microscope (Nikon, Japan) at 520–610 nm wavelength.
Protein extraction and western blotting analysis
When the mice reached 128 days of age, the spinal cord was extracted and homogenized with a protein extraction kit (PRO-PREP; iNtRON Biotechnology, Korea). After homogenization, the extracted samples were centrifuged at 16,000 × g for 10 min at 4℃. Cells that had been treated with several concentrations of KCHO-1 and activated by 10 ng/mL LPS (Sigma) and 10 ng/mL IFN-γ (PeproTech) were lysed with the protein extraction kit (PRO-PREP; iNtRON Biotechnology). All protein samples were quantified by using a Bio-Rad DC protein assay kit (Bio-Rad Laboratories, USA). For western blotting, equal amounts of each sample were loaded and electrophoresed on 10% to 12% SDS-polyacrylamide gels and then transferred to nitrocellulose membranes. The blots were blocked with 5% bovine serum albumin in TBST for 1 h at room temperature and subsequently incubated with various primary antibodies overnight at 4℃. The following primary antibodies were used: beta-actin (sc-47778; Santa Cruz Biotechnology, USA), GAPDH (No. 2118S; Cell Signaling, USA), anti-GFAP (ab7260; Abcam, UK), anti-iNOS (ab15323; Abcam), anti-gp91phox (ab80508, Abcam), anti p-p38 (No. 9211s; Cell Signaling), and p-ERK1/2 (No. 9121; Cell Signaling). The blots were incubated with peroxidase-conjugated secondary antibodies (1:1000; Invitrogen, USA) and developed with enhanced chemiluminescence reagents (Amersham Pharmacia Biotech, USA). Quantification of all immunoblotting data was performed by using ImageJ v. 1.48 software (National Institutes of Health, USA).
Tissue processing and Immunohistochemistry
At 128 days of mouse age, the mice were transcardially perfused with normal saline followed by 4% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) for fixation. Spinal cord portions (L2-L4) were removed, post-fixed in 4% PFA overnight at 4℃, and transferred to 30% sucrose. The tissues were then embedded into a mold filled with optimum cutting temperature (OCT) compound (Tissue-Tek O.C.T Compound; Sakura Finetek, Japan) and stored at −80℃ before cryosectioning. The spinal cords were serially cut on a cryostat into 10 µm thick cross-sections and washed in PBS to remove the OCT compound. After the sections were blocked with 3% to 5% normal goat serum in PBS at room temperature, they were incubated with the primary antibody overnight at 4℃. For detection of the primary antibody, sections were incubated with the appropriate Alexa 488- or 594-conjugated secondary antibodies (1:2,000; Molecular Probes, USA) for 1 h in the dark at room temperature. Images were captured with an Eclipse TE200 confocal microscope (Nikon). To determine the relative immunohistochemical intensity of each signal, ten spinal cord sections per mouse were used (n = 4–5 per group). Each signal was measured in equivalent areas in the anterior horn of the lumbar spinal cord by using ImageJ v. 1.63 software (National Institutes of Health).
Histology
For hematoxylin and eosin (H&E) staining, cryotissues were sectioned at 7 to 8 µm thickness and then stained with H&E by using a standard protocol [9]. The detection of Nissl bodies in the cytoplasm of neurons was performed by Nissl staining. Briefly, the tissues were incubated in 1:1 alcohol/chloroform solution overnight and then immersed in 100% alcohol followed by 95% alcohol and dH2O for rehydration. The rehydrated tissues were stained with 0.1% cresyl violet (Sigma-Aldrich, Germany), rinsed, and then slide mounted with coverslips. Fluoro-Jade C (FJC) staining was used to identify degenerating neurons in the CNS. First, the tissues were immersed in 100% alcohol and then 70% alcohol for 3 min each. The tissues were then transferred to 0.06% KMnO4 for 15 min, washed with dH2O, and stained with a 0.001% FJC solution for 30 min in a dark room. After staining, the slides were rinsed again and mounted with coverslips. Counting Nissl stained or FJC stained motor neurons was performed in equivalent size frames in an anterior horn region, and ten samples were counted per mouse (n = 4–5 per group). All histological examinations were performed by using an Eclipse TE200 confocal microscope (Nikon).
Statistical analysis
Data are presented as mean ± SEM values and were analyzed by using GraphPad 5.0 (GraphPad Software, USA). The immunoblotting data for wild-type, vehicle, and KCHO-1 groups were compared by performing one-way ANOVA. The mean age ± SD of the hSOD1G93A Tg mouse groups were compared by using the Mann-Whitney U test for comparisons between two groups. The log-rank test was used for the Kaplan-Meier survival analyses. Comparisons of Griess assay or ROS concentration results were performed by using the Newman-Keuls post-hoc test of one-way ANOVA results.
Results
KCHO-1 improves motor activity and delays disease onset in symptomatic hSOD1G93A Tg mice
In these experiments, the rotarod test was performed every second day from post-natal day 91 (panel A in Fig. 1), and three or four mice in each group were randomly sacrificed on day 128, which is the average end-stage day for hSOD1G93A Tg mice. Disease onset was determined when mice first fell from the apparatus during a rotarod test, and the average day of disease onset in the vehicle-treated group was day 118.9 ± 1.1, whereas mean onset in the KCHO-1-treated group was delayed until day 127.8 ± 3.1 (panel B in Fig. 1). These results demonstrate that KCHO-1 treatment could significantly delay disease onset of ALS. As shown in panel C in Fig. 1, the rotarod test results indicate the progression of the disease with KCHO-1-treated mice exhibiting a gradual score decrease, while the rotarod scores of the vehicle-treated group rapidly decreased. In particular, between day 114 and day 130, there were significant differences in the rotarod scores of the two groups. The results demonstrate that KCHO-1-treated mice can withstand decreased motor activity during disease progression. To examine the rates of survival between the two groups, the Kaplan-Meier survival test was performed. In the experiment, the KCHO-1-treated group survived longer than vehicle-treated group. The median survival of the KCHO-1-treated group was 139 days while that for the vehicle-treated group was 128 days (panels D and E in Fig. 1). These results demonstrate that KCHO-1 administration in ALS mice can alleviate disease progression and improve survival rate.
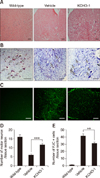
KCHO-1 prevents the degeneration of neurons in the spinal cord of symptomatic hSOD1G93A transgenic mice
To determine whether KCHO-1 administration in ALS mice prevents the degeneration of neuronal cells in the spinal cord, we undertook histological examinations of the anterior horn of the lumbar spinal cord in wild-type, vehicle-, and KCHO-1-treated mice that had been sacrificed on day 128. The spinal cord tissue sections from the three groups were compared by using three different stains. In the first histological examination, H&E staining of spinal cord sections was performed, which indicated that neuronal cell bodies in KCHO-1-treated mice were more numerous than those in the vehicle-treated mice (panel A in Fig. 2). In the second histological examination, we performed Nissl staining of spinal cord sections to identify the Nissl bodies in neurons. The mean numbers of Nissl bodies in the wild-type, vehicle-, and KCHO-1-treated mice were 16.0 ± 1.2, 5.9 ± 0.6, and 11.7 ± 0.6, respectively (panels B and D in Fig. 2). Those staining results indicate that the number of Nissl body-stained neurons in KCHO-1-treated mice was two-fold greater than that in the vehicle-treated mice. Finally, we performed FJC staining, which is used to visualize degenerating neurons via fluorescence microscopy. The FJC stain results for the anterior horn of the spinal cord indicated that the number of degenerating neurons was significantly reduced from 41.5 ± 2.8 per tissue section in the vehicle-treated group to 31.2 ± 1.7 per tissue section after KCHO-1 administration (panels C and E in Fig. 2).
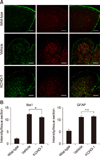
KCHO-1 reduces microglial proliferation and activation
To determine the effect of KCHO-1 administration on microglia and astrocyte distribution in the spinal cord, expression patterns of Iba-1 and glial fibrillary acidic protein (GFAP) were analyzed (panel A in Fig. 3). In the spinal cord, the intensity of Iba-1 expression in microglia was significantly reduced in the spinal cord of the KCHO-1 group (mean ± SEM; wild-type 2.15 ± 0.3, vehicle 12.32 ± 0.39, and KCHO-1 11.05 ± 0.36), whereas the intensity of GFAP expression, a marker indicative of astrocytes, was not significantly different among the groups (wild-type 5.35 ± 0.6, vehicle 11.05 ± 0.57, and KCHO-1 11.39 ± 0.34) (panel B in Fig. 3). The GFAP expression level was also analyzed by using western blot analysis, the results of which did not show any significant differences among the experimental groups (wild-type 0.5 ± 0.1, vehicle 1.51 ± 0.3, and KCHO-1 1.19 ± 0.2) (Supplementary Fig. 1).
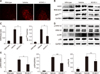
KCHO-1 reduces oxidative stress through gp91phox and MAPK signaling
To investigate the modulation of oxidative stress by KCHO-1 through gp91phox and the MAPK pathway in ALS spinal cord, we first examined the ROS level in the spinal cords of hSOD1G93A Tg mice. As shown in panel A in Fig. 4, the ROS level was reduced in KCHO-1 treated group from that in the vehicle group. Next, we confirmed that the protein levels of inducible nitric oxide synthase (iNOS) and NADPH oxidase (gp91phox) in the spinal cords were readily reduced. The reductions from vehicle-treated to KCHO-1-treated mice were 50% for iNOS (mean ± SEM; wild-type 0.1 ± 0.01, vehicle 0.48 ± 0.04, and KCHO-1 0.25 ± 0.1) and 40% for gp91phox (wild-type 0.23 ± 0.12, vehicle 2.02 ± 0.24, and KCHO-1 1.08 ± 0.13) (panels B and C in Fig. 4). To ascertain that the MAPK pathway was also regulated by KCHO-1, the ERK1/2 and p38 protein levels were examined in the spinal cords. The phosphorylated ERK1 (wild-type 0.28 ± 0.07, vehicle 2.72 ± 0.21, and KCHO-1 1.66 ± 0.31) and ERK2 (wild-type 0.11, vehicle 2.06 ± 0.05, and KCHO-1 1.19 ± 0.14) were significantly reduced by KCHO-1 treatment, but the phosphorylated p38 protein level (wild-type 0.28 ± 0.16, vehicle 1.72 ± 0.11, and KCHO-1 1.19 ± 0.23) was not significantly reduced in the KCHO-1-treated group from that in the vehicle-treated group (panels D and E in Fig. 4).
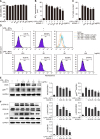
KCHO-1 reduced oxidative stress in BV2 microglial cells
In order to confirm the effect of KCHO-1 on microglia, we activated BV2 microglial cells by administering 10 ng/mL IFN-γ and 10 ng/mL LPS along with varying concentrations of KCHO-1. The KCHO-1 treatment did not affect cell viability until the 100 µg/mL concentration treatment (panel A in Fig. 5). For analysis of the oxidative level in BV2 microglial cells, we performed Griess and ROS level assays. Activation of microglial cells significantly upregulated the NO2− level (mean ± SEM; 17.35 ± 1.27) but KCHO-1 significantly reduced the NO2− level at concentrations of 50 µg/mL (12.17 ± 1.65) and 100 µg/mL (10.65 ± 0.97) (panel B in Fig. 5). We also measured the ROS level in activated BV2 microglial cells with or without KCHO-1. The ROS level was significantly increased following treatment of LPS and the IFN-γ in BV2 microglial cells (48.98 ± 3.8) compared to that in the negative control (5.84 ± 0.7), whereas KCHO-1 treatment at 50 and 100 µg/mL concentrations significantly reduced ROS levels (36.74 ± 1.7 and 37.61 ± 1.2, respectively) (panels C and D in Fig. 5).
At several concentrations, KCHO-1 also reduced the iNOS level and the gp91phox protein level in the activated BV2 microglial cells. Notably, the 100 µg/mL of KCHO-1 significantly reduced iNOS (0.65 ± 0.12) and gp91phox (0.6 ± 0.11) levels from those of activated BV2 microglial cells (iNOS 1.17 ± 0.09, gp91phox 1.32 ± 0.19) (panel E in Fig. 5). To determine the effect of KCHO-1 on the MAPK pathway, we investigated the phosphorylation of p38 and ERK1/2. The LPS and IFN-γ treatment of BV2 cells elevated the phosphorylation of p38 (1.41 ± 0.24) and ERK1/2 (ERK1 1.64 ± 0.24, ERK2 1.33 ± 0.14) from the levels in the negative control (p38 0.07 ± 0.07, ERK2 0.15 ± 0.15). In particular, KCHO-1 at a 100 µg/mL concentration significantly reduced p38 (0.4 ± 0.18) and ERK1 (0.43 ± 0.16) phosphorylation in the activated BV2 microglial cells. KCHO-1 at the same concentration also reduced expression of phosphorylated ERK2 (0.74 ± 0.04), but the reduction was not significant (panel F in Fig. 5).
Discussion
Several previous studies using the herbs included in the compound KCHO-1 have demonstrated that C. longa, C. sinensis, and P. tenuifolia have antioxidant activity [181933], while S. miltiorrhiza, G. elata, and A. japonica have anti-inflammatory or neuroprotective activity [12151625]. In addition, P. japonica and G. uralensis prevent fatigue and have antibacterial effects, and A. carmichaeli can reduce pain [113537]. These herbs have been used for a long time in traditional medicine and have been studied for the treatment of many diseases [1011121516181925333537]. In this study, we found that motor function and survival rate improved and disease onset delayed in KCHO-1-treated hSOD1G93A Tg mice. Furthermore, KCHO-1 administration reduced neuronal loss in the spinal cord of hSOD1G93A Tg mice. Because oxidative stress is a main pathological hallmark of ALS, we focused on the effect of KCHO-1 on reducing oxidative stress [1]. To determine the antioxidant effect of KCHO-1, we studied glial cells, particularly microglia and astrocytes, which play an important role in neurodegeneration. The expression level of Iba-1 was reduced in the spinal cords of the KCHO-1-treated group, whereas expression of the astrocyte marker GFAP was not reduced. These results indicated that KCHO-1 can reduce proliferation of microglia but not astrocytes. Astrocytes are a major source of anti- and pro-inflammatory cytokines, and they contribute to the immune response in the CNS. Previously, astrogliosis, activation, and especially upregulation of GFAP have been observed in the spinal cords of ALS patients. However, the mechanism of astrogliosis in ALS remains unclear. Our data showed that KCHO-1 administration improved motor function, survival rate, and disease onset in hSOD1G93A Tg mice without down-regulating GFAP. Microglia, another type of glial cell, are the immune cells of the CNS, and abnormal activation of these cells influences oxidative stress that regulates the pathological environment in ALS [2039]. In previous studies, the gp91phox subunit of NADPH oxidase was stimulated by mutant SOD1 protein in the microglia of hSOD1G93A Tg mice, and these cells over-produce ROS through NADPH oxidase [2436]. Overexpressed gp91phox protein is a main cause of ROS, but that alone does not have a significant effect on neuronal death. When NADPH oxidase is expressed with iNOS in microglia, the simultaneous activation releases many neurotoxic factors and contributes significantly to oxidative stress [72734]. On that basis, we examined whether the expression of gp91phox and iNOS protein were decreased in the spinal cords of hSOD1G93A Tg mice following KCHO-1 treatment, and our data showed that KCHO-1 reduced oxidative stress in the spinal cord through gp91phox and iNOS down-regulation.
Furthermore, gp91phox-increased oxidative stress in the spinal cord may activate the MAPK pathway, which is known to be a cell stress signaling pathway in the CNS of ALS [1]. In this study, we investigated p38 and ERK1/2 activity and observed that KCHO-1 at several concentrations could alleviate activation of p38 and ERK1/2 in classically activated microglial cells, along with gp91phox and iNOS down-regulation. Consistently, in the hSOD1G93A Tg mice, the phosphorylated form of ERK1/2 was decreased in the KCHO-1 treated group, but the phosphorylated p38 level was not significantly reduced in the KCHO-1 treated group compared to that in the vehicle group. The p38 protein, as a downstream effector of a stress signaling pathway, is activated by various factors in ALS. Particularly, the p38-MAPK pathway can be activated by mutant SOD1 proteins, other inflammatory factors, and by oxidative stress in the astrocytes of ALS. The insignificant downregulation of p38 in the KCHO-1 treatment group may be associated with the lack of a significant effect on astrocytes in the spinal cords of the KCHO-1 group [317]. Therefore, further study is required to elucidate the relationship between the p38-MAPK pathway and the KCHO-1 effect in ALS.
In previous study, it was demonstrated that KCHO-1 can alleviate inflammation via the nuclear factor kappa B pathway in BV2 microglial cells [23] and reduce oxidative stress via HO-1 upregulation [23]. In the present study, we observed that KCHO-1 can alleviate oxidative stress through gp91phox and the MAPK pathway in classically activated microglial cells. Moreover, we have shown that these effects can be elicited in hSOD1G93A Tg mice, thereby improving the progression of ALS. Therefore, KCHO-1 can be considered as a new agent for treatment of ALS.


 XML Download
XML Download