

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Articular pathologies such osteoarthritis (OA) are common diseases in both equines and humans, greatly impacting the daily lives of afflicted individuals and acting as a major cause of wastage in the equine industry. Horses have a double role in joint pathologies because they commonly suffer from these diseases [32], and they are considered the most suitable animal model for testing cell based therapies for human joint injuries [3]. Osteoarthritis is a complex condition characterized by damage to the articular cartilage. Inflammation plays an important role in progression of the disease as the secretion of proinflammatory mediators accelerates cartilage degradation. Conventional treatments primarily focus on relieving this inflammation and controlling pain [11]. An ideal therapeutic approach should stop progressive loss of cartilage and stimulate the regeneration of damaged structures. Treatments for equine joint diseases based on the intra-articular (IA) administration of mesenchymal stem cells (MSCs) are gaining importance because of their regenerative role [8]. MSCs show significant potential for cartilage repair, which is attributed to their trophic and differentiation properties, as well as their immunoregulatory ability [21]. The expression of immunoregulatory molecules by MSCs is regulated by proinflammatory cytokines such interferon-γ (IFN-γ), particularly in combination with tumor necrosis factor-α (TNF-α) [29], which suggests that MSC full immunoregulatory function depends on their activation after inflammatory exposure. Therefore, MSC priming with proinflammatory cytokines prior to their use in vivo might enhance their therapeutic efficacy [5].
Joint injuries result in a variable release of different inflammatory molecules into the synovial fluid. Inflammatory synovial fluid may affect the function and characteristics of IA administered MSCs, such as migratory function or differentiation potential [16], or act as inductors of MSC-immune regulation in equine and human species [1837]. However, the stimulus exerted by synovial fluid could be heterogeneous because variation exists between patients, even within one disease group [18]. Therefore, the combination of definite doses of INFγ with TNF-α leads to a more homogenous stimulus for inducing MSC immunomodulation, which could improve their therapeutic potential. However, the regenerative potential of MSCs through differentiation could be decreased by inflammatory exposure [71720]. In addition, IFN-γ and TNF-α synergistically impair MSC self-renewal, decreasing their effectiveness [38]. To the best of our knowledge, the effects of these cytokines on these properties in equine MSCs have not yet been elucidated.
Because pre-existing joint inflammation may alter the therapeutic efficacy of MSCs when they are IA administered [31], further knowledge of the changes induced in equine MSCs by an inflammatory environment is necessary. Conversely, the use of IFN-γ and TNF-α priming without detriment of viability and plasticity is of major interest to improve cellular therapies. Thus, this study was conducted to assess the possible effects of different inflammatory stimuli on viability and differentiation potential of equine bone marrow derived MSCs (eBM-MSCs) as a step prior for their clinical use in joint injured patients.
Materials and Methods
Experimental design
Equine BM-MSCs (n = 3) were exposed to three different inflammatory conditions and their responses were analyzed: Experiment 1 (SF), 20% allogeneic inflammatory synovial fluid was added to the culture medium to mimic the joint inflammatory environment; Experiment 2 (CK50), IFN-γ (50 ng/mL) and TNF-α (50 ng/mL) were added to the culture medium; and Experiment 3 (CK20), IFN-γ (20 ng/mL) and TNF-α (20 ng/mL) were added to the culture medium. All inflammatory conditions were maintained for 72 h. Subsequently, a proliferation assay was conducted for 7 days. Expression of proliferation and apoptosis related genes was analyzed by real time quantitative polymerase chain reaction (RT-qPCR). Tri-lineage differentiation potential was examined through specific staining and gene expression by RT-qPCR. Triplicate controls were run in each experiment using the same eBM-MSCs (n = 3) cultured with control media.
Animals
Four geldings, named as EQ01, EQ02, EQ03 and EQ04 (weight, 450–500 kg; age, 6–12 years), were used in this study. The horses were patients from the Veterinary Hospital of the University of Zaragoza suffering from varying types of locomotor system injuries, but determined to be in systemic good health based on clinical and hematologic examination. Biological samples (bone marrow [BM] and synovial fluid) were obtained with owner consent and according to local animal welfare regulations.
All procedures were carried out within the Project License 31/11 approved by the in-house Ethic Committee for Animal Experiments of the University of Zaragoza. The care and use of animals were in accordance with the Spanish Policy for Animal Protection RD53/2013, which meets the European Union Directive 2010/63 on the protection of animals used for experimental and other scientific purposes.
Harvesting and characterization of eBM-MSCS
A total of 40 mL of BM from the sternum were collected in heparinized syringes using a 11 g × 101 mm Jamshidi needle (CareFusion, USA) from three donors (EQ01, EQ02 and EQ03). Equine BM-MSCs were isolated using a gradient density separation technique as previously described by our group [27]. The cells were plated and cultured in basal culture medium consisting of low glucose Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 1% glutamine, 1% streptomycin/penicillin and 10% fetal bovine serum (FBS) (all from Sigma-Aldrich, USA) [27]. Cells were expanded until the third passage, then characterized by phenotype determination (flow cytometry and gene expression) and tri-lineage differentiation as previously described [27]. Cells were subsequently frozen in 10% DMSO-90% FBS (Sigma-Aldrich) frozen medium and cryopreserved until experiments started.
Inflammatory synovial fluid collection
Synovial fluid was harvested from EQ04, which presented aseptic synovitis in one tarso-crural joint. Arthrocentesis was performed to reduce the joint effusion and aspirated synovial fluid was collected in heparin-treated tubes (Becton, Dickinson and Company, USA) and used for Experiment 1. Synovial fluid inflammatory status was verified by measuring total protein (g/dL) with a portable optical refractometer (RHB-32 Hand-held Brix Refractometer; Spectrum Technologies, USA), by conducting a cytological examination and determine the concentration of the acute phase proteins (APPs) serum amyloid A (SAA) and haptoglobin (Hp) [2]. Subsequently, synovial fluid was centrifuged at 3,000 × g for 15 min, after which the supernatant was stored at −80℃. All processes were performed under aseptic conditions to prevent culture contamination.
Equine BM-MSCs culture under inflammatory condi
The cytokines used for CK priming were selected based on their described synergy [30]. Synovial fluid and CK inflammatory conditions were determined according to previous reports [29353739]. The time of exposure was determined to be 72 h [26] for the three conditions. For Experiment 1, inflammatory synovial fluid was added to the basal culture medium at 20% (SF medium). For Experiments 2 and 3, basal media were supplemented with recombinant equine TNF-α and IFN-γ (R&D Systems, USA) by adding 50 ng/mL of each cytokine in Experiment 2 (CK50 medium), whereas medium was supplemented with 20 ng/mL of each cytokine for Experiment 3 (CK20 medium). Basal medium was used in control eBM-MSCs in the three experiments (control medium).
One million eBM-MSCs at third passage from EQ01, EQ02 and EQ03 were thawed at 37℃ and seeded at 5,000 cells/cm2 in basal medium at 37℃ and 5% CO2 until confluence reached 80 to 90%, allowing readjustments of conditions prior to initiating the experiments. SF-, CK50-, CK20- or control-media were then added to the cells. Inflammatory exposure and their controls were carried out in triplicate for each animal in each experiment (SF, CK50, and CK20). Cells were cultured for 72 h in their corresponding media at 37℃ and 5% CO2, after which eBM-MSCs were detached with 0.25% trypsin-EDTA (Sigma-Aldrich), washed three times with PBS (Gibco, USA) to completely remove inflammatory molecules, and then used for the different assays.
Proliferation assay
Cell proliferation after every inflammatory exposure and their controls was evaluated over seven days by MTT proliferation assay as previously described [28]. Viable cell numbers for the different samples were determined using a calibration curve consisting of nine triplicate points of increasing amounts of cells [28]. Basal medium was used as a blank. The cell doubling time for the control and inflammatory-stimulated eBM-MSCs from each experiment was calculated.
Tri-lineage differentiation assay
Control and inflammatory exposed cells were cultured in induction medium (differentiation) and basal medium (control) in triplicate according to each differentiation assay. For osteogenic differentiation, eBM-MSCs from each sample were seeded at 20,000 cells/cm2 in 24-well plates. Osteogenic medium consisted of 10 nmol/L dexamethasone, 10 mmol/L β-glycerophosphate and 100 µmol/L ascorbate-2-phosphate (all from Sigma-Aldrich) supplemented basal medium. After 7 days, typical calcium deposit formation was assessed by Alizarin Red staining. To accomplish this, cells were fixed with 70% ethanol for 1 h at room temperature (RT), stained with 2% Alizarin Red stain (pH 4.6) (Sigma-Aldrich) for 10 min (RT) and washed with PBS.
Equine BM-MSCs from each sample were seeded at 5,000 cells/cm2 in 12-well plates for adipogenic differentiation. Adipogenic medium consisted of 1 µmol/L dexamethasone, 500 µmol/L 3-isobutyl-1-methylxanthine, 200 µmol/L indomethacin and 15% rabbit serum (all from Sigma-Aldrich) supplemented basal medium. After 15 days, typical fat droplets were evaluated by Oil Red O staining. Briefly, cells were fixed with 10% formalin (Sigma-Aldrich) for 15 min at RT, stained with 0.3% Oil Red O (Sigma-Aldrich) (dissolved in 60:40; isopropanol: distilled water) stain for 30 min at 37℃, then washed with distilled water.
Chondrogenic differentiation was conducted in pellet culture. To accomplish this, approximately 300,000 eBM-MSCs from each sample were transferred to conic bottom 15 mL tubes, after which 400 µL of chondrogenic differentiation medium were added and the samples were centrifuged at 300 × g for 5 min to pellet the cells. Chondrogenic medium consisted of 10% FBS, 10 ng/mL TGFβ-3 (R&D Systems), ITS+ premix (Becton, Dickinson and Commpany), 40 µg/mL proline, 50 µg/mL ascorbate-2-phosphate, and 0.1 µmol/L dexamethasone supplemented high glucose DMEM (all from Sigma-Aldrich). After 21 days, chondrogenic differentiation was evaluated by specific staining with Alcian blue staining. Pellets were fixed in 10% formalin, embedded in paraffin and sectioned into 5 µm sections. The sections were then hydrated with increasing gradients of alcohol, stained with Mayer's haematoxylin and 3% Alcian blue, rinsed with distilled water, dehydrated with decreasing amounts of alcohols and mounted.
Real time quantitative polymerase chain reaction (RT-qPCR)
Expression of genes coding for molecules related to proliferation (cyclooxygenase 1 [COX-1] COX-1, Cyclin D2) and apoptosis (BAX, BCL-2, BCL-XL, CASP-8, HSP-27, TNF-α, IFN-γ) was assessed by RT-qPCR to investigate eBM-MSCs viability. A RNAspin Mini RNA Isolation Kit (GE Healthcare, UK) was used to isolate total mRNA from approximately 106 eBM-MSCs from each sample. Genomic DNA was removed using DNAse Turbo (Ambion, USA) and 1.5 µg of mRNA from each sample were retrotranscripted to cDNA with a Superscript Reverse Transcriptase Kit (Life Technologies, USA). mRNA isolation and cDNA retrotranscription from osteogenic and adipogenic differentiation assays were performed using a Cells-to-cDNA II kit (Ambion). mRNA could not be isolated from differentiated CK20-eBM-MSCs or from cells undergoing chondrogenic differentiation because there was not enough sample to perform both staining and mRNA isolation. All processes were conducted according to manufacturer's instructions.
A StepOne Real Time PCR System device (Applied Biosystems, USA) was used to perform and monitor RT-qPCR. All reactions were carried out in a total volume of 10 µL with 2 µL of cDNA as the template and Fast SYBR Green Master Mix (Applied Biosystems). Amplification and analysis were performed as previously described [27]. The expression levels of all genes in each sample were normalized by a normalization factor (NF), which was calculated as the geometric mean of the quantity of two housekeeping genes, GAPDH and B2M [15]. The Primer Express 2.0 software (Applied Biosystems) was used to design primers based on known equine sequences. Information about primers is shown in Table 1.
Statistical analysis
Data obtained in this study were subjected to statistical analysis using SPSS 15.0 (SPSS, USA). Proliferation data are presented as the means (n = 3) ± SEM cell count of stimulated and unstimulated eBM-MSCs at each time point (7 days) for each experiment. Differences in mean cell numbers between stimulated and unstimulated eBM-MSCs from each day were analyzed by paired Student's t tests separately for each experiment. RT-qPCR data are reported as the means (n = 3) ± SEM fold increase or decrease of stimulated eBM-MSCs gene expression over unstimulated control eBM-MSC. Differences between eBM-MSCs from every inflammatory condition (SF, CK50 and CK20) and their controls were analyzed by paired Student's t-tests. The significance level was set at p < 0.05 for all analyses.
Results
Isolation and characterization of eBM-MSCS
Cells obtained from EQ01, EQ02 and EQ03 BM aspirates showed plastic-adherent fibroblast-like morphology and were positive for the surface markers CD90, CD105, CD44 and MHC-I and negative for MHC-II by flow cytometry. All samples expressed transcripts for CD105, CD90, CD73, CD44 and MHC-I, but not for CD34 and CD45, and low levels for MHC-II by RT-qPCR. The ability of tri-lineage was confirmed in all cases (data not shown).
Inflammatory synovial fluid harvesting
The average total protein measurement in the harvested synovial fluid was 2.25 g/dL. The SAA concentration was 0.585 × 10−3 g/dL and the Hp concentration was 0.023 g/dL. These data confirmed the inflammatory status of the synovial fluid, and when combined with the cytological examination discarded a septic origin of the joint inflammation.
Effects of inflammatory environment on eBM-MSCs proliferation
For the MTT assay, the total number of viable cells from every sample was determined by extrapolation from the calibration curve: y = 9 × 10−6 x + 0.0805, r2 = 0,939, where y represents the optical density of the well and x represents the amount of cells. Synovial fluid-exposed eBM-MSCs grew in a similar manner to control cells (panel A in Fig. 1). In both CK50 and CK20 experiments, the number of CK-stimulated cells decreased until the fourth day, then increased slightly until the sixth day, but still remained below the initial level (panels B and C in Fig. 1). Significant differences (p < 0.05) in the number of cells were observed between control and CK-exposed cells in all days under both CK20 and CK50 conditions. As shown in Table 2, the cell doubling times were also similar between control and SF stimulated eBM-MSCs, but increased greatly in both CK50 and CK20-stimulated eBM-MSCs. The proliferation related genes COX-1 and Cyclin D2 were significantly downregulated under the CK50 and CK20 conditions (p < 0.05), and their expression remained similar in the SF experiment (Fig. 2).
Effect of inflammatory environment on apoptosis gene expression of eBM-MSCs
Apoptosis-related gene expression was studied in CK50 and CK20 exposed eBM-MSCs. BAX was significantly downregulated under both CK50 (p < 0.01) and CK20 (p < 0.05) conditions, while BCL-2 was only significantly downregulated under CK20 conditions. CASP8 showed a trend to increase its expression under both CK conditions. The anti-apoptotic factor heat shock protein 27 (HSP27) was significantly downregulated under CK50 conditions (panel A in Fig. 3). TNF-α expression was downregulated and no-expression was found for IFN-γ under SF conditions, whereas significant upregulation of TNF-α (p < 0.05) and IFN-γ (p < 0.05) was observed in CK50 and CK20, respectively (panel B in Fig. 3).
Effect of inflammatory environment on eBM-MSCs differentiation potential
Osteogenic, adipogenic and chondrogenic differentiation were achieved in control (unstimulated) cells from the three experiments. Spontaneous differentiation was not observed in any of the non-differentiated controls.
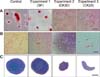
Osteogenic differentiation was confirmed in SF-exposed eBM-MSCs by positive staining of the calcium deposits with Alizarin red. CK-stimulated eBM-MSCs showed irregular morphology, but did not form a calcium-rich matrix (panel A in Fig. 4). Alkaline phosphatase (ALP) and Runt-related transcription factor 2 (RUNX2) gene expression was not significantly modified by SF conditions, but the expression of both genes in SF-exposed eBM-MSCs was slightly higher than in the differentiated control (panel A in Fig. 5). Expression of ALP and RUNX2 was downregulated in CK50-exposed eBM-MSCs compared to the differentiated control; however, this downregulation was only significant for RUNX2 (p < 0.05) (panel B in Fig. 5). ALP increased by 18.93-fold in SF-differentiated cells relative to the control, but only by 2.49 in CK50-differentiated cells. RUNX2 expression in SF-differentiated cells increased by 4.48-fold, whereas a 3.85-fold decrease was observed in CK50-differentiated cells (panels A and B in Fig. 5).
Oil red O staining revealed lipid droplets in SF-exposed eBM-MSCs undergoing adipogenic induction. Both CK50 and CK20-stimulated cells did not display an adipogenic phenotype (panel B in Fig. 4). Peroxisome proliferator-activated receptor γ (PPARγ) and lipoprotein lipase (LPL) mRNA levels were not significantly modified by SF exposure (panel C in Fig. 5). LPL and PPARγ in CK50- and SF-differentiated cells showed similar increases relative to non-differentiated control cells, but both genes were significantly downregulated in CK50-exposed cells (p <0.05) compared to unstimulated differentiated cells (panel D in Fig. 5).
Synovial fluid-exposed cells maintained their chondrogenic differentiation potential, as indicated by the positive blue staining of the extracellular matrix upon Alcian blue staining. However, neither CK50 nor CK20-exposed eBM-MSCs differentiated into the chondrogenic lineage, and they formed irregular discoid pellets instead of the typical spherical shape (panel C in Fig. 4).
Discussion
Both APPs analyzed in the inflammatory synovial fluid were above the reference ranges [114], which is in agreement with the expected changes for these proteins [14]. The inflammatory environment provided by cytokine priming resulted in a remarkable viability impairment of eBM-MSCs, whereas SF conditions did not alter their regular growth. The significant down-regulation of COX1 and CyclinD2, enzymes involved in the regulation of the cell cycle and cell proliferation [3336], in both CK experiments might be related to the failed proliferation of eBM-MSCs [3336]. Thus, treatment with both cytokine concentrations appeared to be cytotoxic. To elucidate this phenomenon under CK conditions, the expression of several genes involved in the apoptosis process was investigated. BCL2-Associated X Protein (BAX) is a proapoptotic gene and BCL-2 is an antiapoptotic gene, both of which are involved in the apoptosis mitochondrial pathway [13]. While BAX was downregulated under both CK conditions, BCL-2 expression showed contrasting results between CK50 and CK20 experiments, which suggest that the apoptosis mitochondrial pathway might not be involved. However, the apoptosis death receptor pathway might be involved since CASP8 showed a trend to upregulation [13]. In addition, the downregulation of HSP27, which protects against injury-related MSC apoptosis [34], might be associated with failed cell development and cell differentiation [34]. TNF-α and IFN-γ have a synergic effect in the induction of apoptosis by the death receptor signaling pathway in MSCs [20]. Additionally, TNF-α converts the signaling of the non-apoptotic receptor Fas, which is activated by IFN-γ, into a CASP8 proapoptotic cascade in MSCs [2038]. Based on these findings, the apoptotic process might be involved in the low viability and plasticity of CK-exposed eBM-MSCs.
Reduction of the tri-lineage differentiation ability of MSCs by proinflammatory cytokines has been described in other species. T-cells mediated secretion of TNF-α and IFN-γ synergistically hamper osteogenesis, chondrogenesis [6923] and adipogenesis [17]. TNF-α or IL-1β exposure significantly reduces the gene expression of osteogenic, adipogenic and chondrogenic markers in murine MSCs [17]. In humans and rats, these cytokines inhibit the expression of the osteoblast differentiation transcription factor, RUNX2 [79]. Moreover, TNF-α plays a dual role in osteogenesis. On the one hand, it inhibits the osteoblast differentiation transcription factor RUNX2 [9]. On the other hand, TNF-α can enhance the MSC osteogenic differentiation in a dose-dependent manner, although this effect is avoided in the presence of anti-inflammatory agents, such as dexamethasone, which is added to the osteogenic-induction media [22]. It has been proposed that the inhibition of osteoblast differentiation by TNF-α occurs through the p55 TNF receptor [10]. High secretion of IFN-γ by immune cells has also been shown to prevent osteogenesis in MSCs in allogeneic implants in mice by dramatically reducing the expression of ALP and RUNX2 [6]. Our results agreed with those of previous reports in others species since the transcript levels of ALP and RUNX2 detected were lower in the presence of TNF-α and IFN-γ Adipogenesis could also be impaired by inflammatory conditions because high levels of proinflammatory cytokines, such as TNF-α, lead to a reduction in PPARγ expression by MSCs [4]. In agreement, PPARγ gene expression was down-regulated under our CK conditions, resulting in a commitment of adipogenic differentiation. TNF-α and IFN-γ have also been described as inhibitors of chondrogenesis and collagen formation in rat and mouse MSCs [1724]. Chondrogenesis has been shown to be hindered through the inhibition of the chondrogenic factor Sox9 caused by these cytokines [23]. According to these reports, our CK conditions also led to a lack of chondrogenic potential in eBM-MSCs. The effects of proinflammatory cytokines on MSC differentiation could vary between species and, to our knowledge, this is the first time that this effect has been studied on equine MSCs. Overall, our results support the findings reported for other species [717] demonstrating the inhibitory effect of proinflammatory cytokines on the tri-lineage differentiation of eBM-MSCs.
The natural joint environment could be favorable for chondrogenic differentiation [1225]. However, when proinflammatory cytokines are present in high amounts, synovial fluid turns into an inflammatory environment, preventing chondrogenesis [19]. Depending on the level of proinflammatory cytokines, inflammatory synovial fluid could be more or less harmful for chondrogenesis and cartilage matrix formation (i.e., rheumatoid arthritis vs. OA) [16]. Under our experimental conditions, tri-lineage differentiation was achieved by SF-exposed eBM-MSCs, suggesting that a moderate inflammatory status of the synovial fluid did not have a significant effect on eBM-MSCs differentiation ability.
The present research provides novel results about eBM-MSCs in inflammatory environments that can contribute to their intra-articular use for therapeutic applications. In summary, an inflammatory synovial environment is satisfactorily tolerated by eBM-MSCs, which maintained their proliferation and differentiation abilities, encouraging the use of cell therapy for joint pathologies. In contrast, cytokine priming tested in this study negatively affected the eBM-MSCs proliferation and differentiation abilities, and appeared to induce apoptosis, possibly compromising their in vivo efficacy and their therapeutic potential.





 XML Download
XML Download