

 Citation
Citation Print
Print


Introduction
Mammary tumors are a common neoplastic disease in female dogs, especially in intact animals and animals spayed after the second heat. Approximately half of canine mammary tumors (CMTs) are considered malignant and these can severely compromise the quality of life of the affected dogs due to local recurrence, distant metastases, and finally death [82227].
CMTs have been demonstrated to share many features with human breast cancers, including histological appearance, hormone-dependence, and biological behavior [202527]. A recent genome-wide comparative analysis of human and CMTs has demonstrated a great degree of similarity in the perturbations of many cancer-related pathways, including the Wnt/β-catenin signaling pathway [28]. The Wnt/β-catenin signaling pathway is vital in embryonic development and tissue self-renewal, and it regulates various processes that are important for cancer progression [219]. With the activation of the Wnt/β-catenin pathway, the Wnt ligand binds to frizzled (FZD) receptors and its co-receptors low-density lipoprotein receptor-related proteins 5/6 (LRP5/6) on the cell membrane, forming a complex after recruitment of Dishevelled. Subsequent LRP6 phosphorylation and recruitment of the Axin complex to the receptors lead to the stabilization of β-catenin, which then accumulates and translocates to the nucleus to form complexes with T-cell factor/lymphoid enhancer-binding factor (TCF/LEF) and activate the expression of Wnt target genes, such as cyclin D1 and matrix metalloproteinase-7 (MMP7), to promote cell growth, proliferation, and differentiation as well as aiding in tumor cell invasion and metastasis [2,1016].
In a recent study performed on cell lines derived from CMTs, the canonical Wnt signaling pathway was shown to be activated with aberrant expression of lymphoid enhancer-binding factor 1 (LEF1) [6]. A few immunohistochemistry studies have also documented an alteration in the expression of protein β-catenin in CMTs in parallel with the acquisition of a more invasive and undifferentiated histological phenotype, but how this affects prognosis is unclear [1424]. Information about the expression profiles of other genes related to the Wnt signaling pathway in tumorigenesis of CMTs is lacking.
In this study, we employed an RT2 Profiler PCR Array to analyze the expression of Wnt signaling components (Wnt ligands, receptors, targets and β-catenin destruction complex) and some of the genes related to Wnt signaling (such as Wnt antagonists dickkopf-1 protein [DKK1] and secreted frizzled-related sequence proteins [SFRPs]) in CMTs. We also investigated the expression of β-catenin and cyclin D1, two critical molecules in canonical Wnt signaling pathway, by using immunohistochemistry and Western blotting assays in CMT samples.
Materials and Methods
Retrospective samples and histopathology
One hundred CMT samples were collected from 100 bitches underwent regional or unilateral mastectomy and ovariohysterectomy in the Veterinary Teaching Hospital of China Agricultural University from September 2012 to October 2014. Normal mammary gland tissues (NMGTs) were collected (at necropsy) from 8 intact healthy dogs that were used for general surgery practice courses for undergraduate students at the College of Veterinary Medicine. These procedures were approved by the Animal Care Committee of China Agricultural University (ID: 1114120800096). Survival of CMT patients was determined by telephoning the dog owners one year after the surgery.
After routine histologic processing and hematoxylin-eosin staining, CMT samples were evaluated by a board-certified pathologist and classified and graded as described by Goldschmidt et al. [5] and Peña et al. [18], respectively. The presence of necrosis and lymphatic invasion in the histological sections was also recorded.
RT2 Profiler PCR Array tests
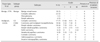
A custom RT2 Profiler PCR Array (CAFP12858; Qiagen, Germany) that could simultaneously detect 30 genes related to the Wnt signaling pathway in canines (Table 1) was used. Each array plate contained 2 sets of 48 wells for 2 tests. Genomic DNA contamination, reverse transcription, and positive PCR controls were included in each 48-well set on each plate. Glyceraldehyde-3-phosphate-dehydrogenase (GAPDH) was used as the assay reference gene.
Thirty-five CMT samples (see Table 2) and 5 NMGT samples were used in the assays. Total RNA was isolated from each frozen tissue sample using TRIzol Reagent (Life Technologies, USA) and quantified with Nanodrop 2000 (Thermo Fisher Scientific, USA). Then total RNA (1 µg) was reverse transcribed in a final volume of 20 µL with an RT2 First Strand Kit (catalog No. 330401; Qiagen). The RT2 Profiler PCR Array tests were performed following the instructions of the manufacturer. Briefly, all cDNA in each sample (20 µL) was first diluted with RNase-free water to a volume of 111 µL. Then 51 µL of the diluted cDNA was mixed with 675 µL 2 × RT2 SYBR Green ROX qPCR Mastermix (catalog No. 330523; Qiagen) and 624 µL RNase-free water. Of that mixture, 25 µL per well was added into the 48 wells of the array plate. The qPCR was carried out using the Applied Biosystems 7500 Real-Time PCR System (software ver. 2.0.6; Applied Biosystems, USA) under the following thermal cycling conditions: 10 min at 95℃, followed by 40 cycles of 15 sec at 95℃ and 1 min at 60℃. The exported Ct values were input to a template Excel file provided by SABiosciences (Qiagen) and uploaded for the online analysis. After data review, qualified data from 35 CMT and 5 NMGT samples were analyzed by applying the 2−ΔΔCt method.
Immunohistochemistry (IHC) evaluation and immunohistochemical scores (IHS)
Paraffin sections of CMT and NMGT samples were dewaxed in xylene and subsequently rehydrated through an ethanol series. Antigen retrieval was carried out by microwave treatment in 0.1 M citrate buffer (pH 6.0) for β-catenin or EDTA buffer (pH 9.0) for cyclin D1. Endogenous peroxidase activity was blocked with 3% hydrogen peroxide for 20 min. Then normal goat serum was applied on the slides to block non-specific binding for 20 min. Rabbit anti-β-catenin polyclonal antibodies (1:5,000 dilution, ab6302; Abcam, USA) or rabbit anti-cyclin D1 polyclonal antibodies (1:1,000 dilution, ab185241; Abcam) were applied on the sections overnight at 4℃. The sections were visualized by using the Polink-2 HRP plus rabbit DAB detection system (Golden Bridge International, USA) and counterstained with hematoxylin. Negative controls in which PBS replaced the primary antibody were set at the same time.
The IHS of β-catenin was evaluated by determining the percentage of positive cells and the intensity of the cytoplasmic staining in accordance with the description of Khramtsov et al. [11]. The scores for both membrane-associated and cytoplasm-associated β-catenin ranged from 0 to 3. The IHS of cyclin D1 was assessed by determining the percentage of stained nuclei. The staining was scored as 0 when the sample had no nuclear staining or if < 10% of its nuclei were stained; whereas tumor samples with > 10% of its nuclei stained were scored as 1 [17]. The percentage of positive staining was estimated by counting no less than 500 cells in 3 randomly selected high-power fields.
Western blotting analysis
Eleven CMT samples (including 4 benign CMTs and 7 malignant CMTs) and 1 NMGT sample were selected for the detection of expressions of β-catenin, TCF7, LEF1, and cyclin D1. Proteins were extracted with RIPA lysis buffer (Macgene, China) and the soluble protein was resolved by performing electrophoresis through 12% SDS-PAGE gels followed by transferring to polyvinylidene difluorid membranes by semi-dry blotting. Non-specific binding sites were blocked by incubation for 1 h in 5% w/v milk powder in PBS, after which membranes were sliced and separately probed with rabbit polyclonal antibodies against β-catenin (diluted at 1:4,000; ab6302; Abcam), LEF1 (diluted at 1:600; ab83964, Abcam), TCF7 (diluted at 1:4,000; ab30961, Abcam), or cyclin D1 (diluted at 1:1,000; ab185241, Abcam). HRP conjugated goat anti-rabbit IgG or goat anti-mouse IgG were used as the secondary antibody. Blots were developed in ECL reagent (Macgene) and exposed to X-ray film.
Statistical analysis
For statistical purposes, Grade I tumors were assigned to the low malignant (LM) group, while Grades II and III tumors were clustered together and assigned to the highly malignant (HM) group. Similarly, some tumor subtypes were grouped together based on similarities in morphology and/or biological behavior, leading to the formation of 4 subtype groups (detailed in Table 2). SPSS (ver. 20; IBM, USA) was used for statistical analysis. A one-way ANOVA with Tukey's post hoc test was performed for analysis of the fold changes of genes in the RT2 Profiler PCR Array test results. A chi-squared test and Fisher's exact test were used for analysis of the associations of β-catenin or cyclin D1 expressions with clinicopathological features. A significant difference was expressed as *p < 0.05 while a highly significant difference was expressed as **p < 0.01.
Results
Histopathological examination of CMT samples
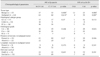
Histopathological examination of the 100 CMT samples revealed 37 benign tumors and 63 malignant tumors, which were distinguished in four subtypes as summarized in Table 2. Among the 100 tumors, 37 were included in subtype group 1, 17 in subtype group 2, 10 in subtype group 3, and 36 in subtype group 4. Among the 63 malignant tumors, 30 were classified as LM and 33 as HM. Necrosis and lymphatic invasion were detected in 41 and 9 malignant cases, respectively. The 8 NMGT samples were confirmed to be non-neoplastic, with only mild hyperplastic changes (lobular hyperplasia) detected in 4 samples.
Evaluation of transcriptional expression of genes related to Wnt signaling in CMTs
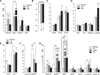
Based on the grading and classification of the CMT samples (Table 2), 9 benign tumors, 26 malignant tumors, and 5 NMGT samples were subjected to RT2 Profiler PCR Array assays. Among the 30 simultaneously detected genes, 9 exhibited significant changes in expression (Fig. 1). Expression of WNT5A was significantly upregulated in LM tumors (panel A in Fig. 1). Expression of DKK1 was correlated with the degree of malignancy, showing a 16-fold change in LM tumors and a 29-fold change in HM tumors (panel A in Fig. 1). In contrast, SFRP4 was significantly downregulated in both LM and HM tumors (7.1-fold and 7.0-fold changes, respectively, panel B in Fig. 1). For β-catenin and the destruction complex-related factors investigated in this study, only β-catenin showed an obviously upregulated expression in HM tumors (6.8-fold increase, panel D in Fig. 1). A significant difference was detected in the expression of transcription factors TCF7 and LEF1 between the benign tumor and the NMGT samples (panel E in Fig. 1). Expression of the Wnt target gene MMP7 was increased in the benign CMTs (20-fold change) while it decreased in LM tumors and HM tumors (9-fold and 6.5-fold changes, respectively; panel F in Fig. 1). The data were reanalyzed after grouping the tumor samples into 4 subtypes, and the results of the analysis showed that, compared with the expressions in other subtypes and NMGTs, expressions of SFRP1 and DKK1 were highest in carcinoma and malignant myoepithelioma (CMM) subtypes. The expression pattern of β-catenin was similar to that of the SFRP1 and DKK1 (panel G in Fig. 1).
Immunohistochemical evaluation of β-catenin and cyclin D1 expression in CMTs
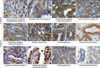
Expressions of β-catenin and cyclin D1 were further evaluated by performing immunohistochemical analysis. Compared to NMGTs (panel H in Fig. 2), significantly reduced membrane expression of β-catenin (IHS as 0 and 1) was observed in all tumor samples (panel A–G in Fig. 2; p = 0.000). Most (8/10) CMMs also showed weak to moderate intensity staining of cytoplasm (panel A in Fig. 2); while moderate to strong cytoplasm staining of β-catenin (IHS as 2 and 3) was detected in the other CMT subtypes (panel B–G in Fig. 2). Expression of β-catenin in the cytoplasm was significantly higher in CMTs than in NMGTs (Table 3). There was an obvious shift of β-catenin localization from membrane to cytosol in the comparison of CMTs with NMGTs (p = 0.000; Fig. 3). Nuclear expression of cyclin D1 (scored as 1) was observed in 84% (84/100) of CMTs (25 benign and 59 malignant tumors; panel I–K in Fig. 2), while its expression was absent or less than 10% (scored as 0) in the nuclei of NMGTs (panel L in Fig. 2), a statistically significant difference (p = 0.000; Table 3). There was significantly higher nuclear expression of cyclin D1 in malignant tumors than in benign tumors (p = 0.001; Table 3). With regard to the expression of β-catenin in cytoplasm and the nuclear staining of cyclin D1, there were no significant differences detected among the different histological grades, between presence/absence of necrosis or lymphatic invasion in malignant CMTs, and between death or survival at one year after surgery (Table 3).
Western blotting analysis of the expression of β-catenin, TCF7, LEF1 and cyclin D1
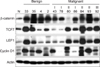
The expression of β-catenin increased significantly in 10 of 11 detected CMTs; the exception, a decreased expression, was detected in a case with simple tubulo-papillary carcinomas. Increased LEF1 expression was detected in complex adenomas, intraductal papillary carcinomas, and solid carcinomas, while no (or slight) increases of LEF1 expression were detected in simple adenomas, complex carcinoma, and 1 simple tubulopapillary carcinomas (Grade I). The expression levels of TCF7 were similar to those of LEF1 in CMTs. Decreased cyclin D1 expression was only detected in a case of simple tubulo-papillary carcinomas (Fig. 4).
Discussion
The RT2 Profiler PCR Array system is a reliable tool for analyzing the expression of multiple genes and thus is extensively used for the analysis of diverse signaling pathways in human and model animals [1215]. In the current study, we employed this tool for the analysis of transcriptional profiles of Wnt signaling components in CMTs. Among the 30 selected genes involved in the Wnt signaling pathway, remarkable expression changes at the mRNA level were observed for several Wnt-related genes (e.g., the upregulation of WNT5A, FZD3, and LEF1 and the downregulation of SFRP4) in different types of CMTs. Based on previous reports [16], the results obtained in this study suggest that Wnt signaling components have important roles in tumorigenesis and tumor progression of CMTs.
Of the activated Wnt signaling components in CMTs, both DKK1 and SFRP1 were significantly upregulated in CMM samples, even in malignant CMTs. This is of particular interest as it may suggest a different tumorigenesis pathway for malignant myoepithelioma, as a previous study showed a significantly high Ki67 expression in such tumors [21]. Further studies using laser capture microdissection to isolate the myoepithelial component of these tumors could help to clarify the present finding. As detection of DKK1 and SFRP1 has been extensively reported following diagnostic studies of liver, pancreas, lung, prostate, and nasopharyngeal cancers of humans [723263031], the present result provides further evidence for the possible use of DKK1 and SFRP1 as valuable candidate biomarkers for identifying malignant CMTs.
By performing IHC assays, cytoplasm accumulation and weak membrane staining of β-catenin was detected in most CMT samples, whereas β-catenin nuclear staining was infrequently observed, indicating that Wnt/β-catenin signaling is aberrant in CMTs and, thus, could be an indicator of this kind of tumor. This phenomenon could be partially explained by the transference of β-catenin from the cell membrane to the cytoplasm along with dissociation of E-cadherin and the β-catenin complex [929]. In addition, we did not detect a correlation between the intensity of β-catenin expression and the survival period of CMT patients (data not shown). Concerning the inconsistencies among previous reports on the correlation between the expression and subcellular location of β-catenin and the prognosis of human breast cancer and CMTs [341314], further studies are required to understand the roles played by β-catenin in the tumorigenesis and tumor progression of CMTs.
In conclusion, this study substantiated activation of the Wnt signaling pathway in CMTs by performing transcriptional profiling of 30 genes in RT2 Profiler PCR Array assay results and determining ectopic subcellular distributions of β-catenin and cyclin D1 by IHC assay results. The most activated Wnt pathway-related genes, such as DKK1 and SFRP1, may have potential as predictive biomarkers of and targeted therapeutics for CMTs.

 XML Download
XML Download