

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Fowl typhoid is caused by Salmonella Gallinarum, the transmission route of which is horizontal, i.e., from an infected ovary to the egg and the progeny [3]. The disease can be either acute or chronic and usually results in a high mortality rate (60%) and increased morbidity [229]. Although Salmonella Gallinarum has adapted to its avian host and rarely induces food poisoning in humans, fowl typhoid outbreaks cause severe economic losses worldwide [36].
The worldwide increase in multi-antibiotic resistance in bacteria is a problem caused by major drug resistant nosocomial pathogens. As a result of multiple antibiotic inefficiencies, the mortality rate caused by Salmonella has increased [12]. Recently, several attempts have been made to prevent salmonellosis by using alternatives to antibiotics, such as vaccination, bacteriophages, probiotics, and herbs [101315]. However, these methods have demonstrated limited efficiency. Therefore, the development of efficacious feed additives to prevent poultry salmonellosis is a major research goal [120].
Formulation of balanced diets is fundamental to economical poultry production, and this process depends on knowledge of the nutrient requirements of poultry and the nutritional attributes of various feed sources. Minerals are naturally occurring inorganic solids and dietary nutrients essential in the maintenance of homeostasis for laying hens [22]. In addition, minerals have important roles in pathogen virulence and host antimicrobial resistance [2734]. Calcium enhances intestinal resistance to infectious diseases and protects against Salmonella- and enterotoxigenic Escherichia coli (ETEC)-associated diarrhea [56]. Iron deficiency is associated with lowering resistance to infection, whereas iron supplements reverse this effect [212326].
Recently, organic minerals (chelated minerals), because of their potential for higher bioavailability than inorganic minerals, have been studied by several researchers [19]. Organic minerals include any mineral bound such that it is chelated or bonded to organic molecules, such as an amino acid, thereby forming chemical structures with unique characteristics such as stability and high mineral bioavailability. Several reports have indicated that organic sources of trace minerals have a higher bioavailability than that of inorganic forms [41937]. After absorption, organic minerals may induce physiological effects, which could improve specific metabolic responses, such as the immune response. Many studies have demonstrated the benefits of metal-amino acid chelates on animal metabolism [33].
In this study, we evaluated the effects of the feed additive guanosine 5′-monophosphate (GMP)-chelated calcium and iron (CaFe-GMP), a metal-nucleotide chelate, against salmonellosis in layers.
Materials and Methods
Preparation of organic mineral feed additives
The CaFe-GMP was provided by Medinutrol (Korea). CaFe-GMP, an organic formulation in which calcium and iron are provided as GMP chelates was used as a supplement to the layers basal diet. A commercial, nutritionally complete, antibiotic-free, general layer feed was used as a basal diet to meet the nutrient requirements for layers in accordance with Nutrient Requirements of Poultry recommendations [22]. The basal diet was formulated to contain a given chemical composition (Table 1).
In our previous study, the potential effect of prebiotics on the growth of Lactobacillus spp. in layers using a feed supplement of 0.16% CaFe-GMP, which contained 83 ppm calcium (Ca) and 55 ppm iron (Fe) (unpublished data) was investigated. In the present experiment, for the CaFe-GMP additive group, CaFe-GMP was completely mixed into the basal diet to form 0.16% CaFe-GMP containing 83 ppm Ca and 55 ppm Fe (Table 1).
Animal experiment
Hy-line brown layer chickens (34 weeks old) with no history of salmonellosis were obtained from a local farm. Microbiological testing did not detect Salmonella Gallinarum in the feces of the chickens. All chickens were maintained at the Chonnam National University College of Veterinary Medicine (Chonnam, Korea) in separate air-controlled rooms. Chickens were allowed free access to tap water, and their respective diets were supplied three times per day. All experimental procedures were approved by the Institutional Animal Care and Use Committee of the Chonnam National University (approval No. CNU IACUC-YB-2015-59).
After a 7-day acclimation period, the chickens were divided into three groups with six chickens in each group. The groups were as follows: normal group, fed with a normal commercial layer feed without Salmonella Gallinarum inoculation; control group, fed with a normal commercial layer feed but with a Salmonella Gallinarum inoculation; additive group, fed with a normal commercial layer feed supplemented by the addition of CaFe-GMP and with Salmonella Gallinarum inoculation. All chickens were fed their respective diet for 4 weeks, and all but the normal group were inoculated with Salmonella Gallinarum. The experiment was conducted in triplicate.
Preparation of Salmonella Gallinarum (inoculum)
Salmonella Gallinarum (SG3001) for experimental inoculation was provided by the Animal and Plant Quarantine Agency (Korea). The SG3001 was cultured on MacConkey agar plates overnight. One colony was selected and inoculated in Brain Heart Infusion (Difco Laboratories, USA) broth and incubated at 10 × g for 6 h at 37℃. Viable counts of the broth culture were obtained by using the spread plate method. Inoculated broth was diluted with phosphate buffer saline (PBS; Sigma-Aldrich, USA) to obtain an infective dose of 1 × 108 CFU/mL (CFU, colony-forming unit) (the optimal dose was determined in preliminary experiments). Experimental inoculations were administered orally.
Body weight, clinical symptoms, and pathological changes
Body weights of chickens were measured at 0 and 5 days post-infection (dpi). The chickens were monitored daily for clinical signs, including anorexia, diarrhea, and depression, among others. Postmortem examinations of all dead and sacrificed chickens were conducted and any pathological changes associated with fowl typhoid were noted. Additionally, the mortality rate in each group during the experimental period was recorded.
Experimental parameters measured of poultry production
Egg production (eggs/chicken/day, %), egg loss (broken and cracked eggs, %), and egg weight (g) were recorded daily. Feed intake (g/chicken/day) was calculated by weighting the initial feed input against the uneaten food over the experimental period. Feed conversion rate (FCR; g feed/g egg) were determined on a daily basis. FCR was expressed as kilograms of feed consumed per kilogram of egg produced. Mortality rate (%) was recorded daily as it occurred.
Re-isolation of Salmonella Gallinarum from internal organs
Re-isolation numbers for Salmonella Gallinarum was determined for liver, spleen, and feces collected from each sacrificed chicken at 5 dpi. In addition, feces were retrieved from sterilized paper that was used to line the chicken's housing unit. For non-selective pre-enrichment processing, 0.1 g of tissue and feces from the cecum of each sample were aseptically collected and added to 10 mL of sterile Buffered Peptone Water (Sigma-Aldrich), homogenized completely, and incubated at 37℃ for 24 h. Then, the homogenate was transferred from 1 mL of the pre-enrichment broth to 10 mL Tetrathionate Broth (TTB; Difco Laboratories) and incubated at 37℃ for 24 h for selective enrichment processing. The TTB was serially diluted 10-fold in PBS, and 50 µL of each dilution was spread onto a Salmonella Shigella agar (Difco Laboratories) plate and incubated at 37℃ for 24 h. The resulting characteristic black-colored colonies were counted and expressed as CFU/0.1 g tissue, but only for those plates with counts of 30 to 300 colonies per plate.
Results
Changes in body weight and clinical symptoms after Salmonella Gallinarum infection
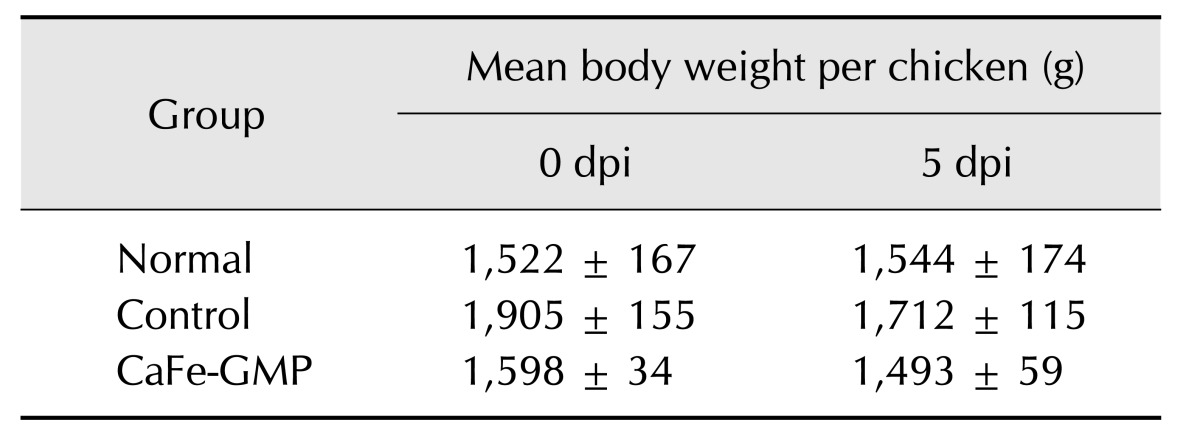
The adverse effects of the Salmonella Gallinarum infection induced in the present study were assessed by examining body weight and clinical symptoms. For each group, body weight change was compared between mean weight at 0 dpi for all chickens in a group to that at 5 dpi in the surviving chickens. The body weights of each group at 0 and 5 dpi are shown in Table 2. The non-inoculated, normal group did not exhibit a change in body weight, whereas the inoculated, control group (1,741.0 g ± 125.0 g) had significantly lower body weight at 5 dpi than at 0 dpi (1,905.7 g ± 155.3 g), a loss of approximately 8.6% (p < 0.07). However, the inoculated, CaFe-GMP additive group exhibited only slightly lower body weight at 5 dpi (1,493.6 g ± 59.4 g) than at 0 dpi (1,598.0 g ± 34.3 g); the change was not significant. The percentage decline in body weight for the CaFe-GMP additive group was less than that of the control group. Clinical signs (anorexia, diarrhea, and depression) were observed in the control group, but the CaFe-GMP additive group showed very few typical clinical signs of Salmonella Gallinarum.
Changes in mortality rate of layers experimentally infected with Salmonella Gallinarum
Mortality rate is a major evaluation marker used in treatment toxicity analysis for preclinical studies. Mortality rates in the three trial groups are summarized in Table 3. Mortality was first observed at 4 dpi in the control group, whereas there was no mortality in the CaFe-GMP additive group or the normal group during the experimental period. Although low levels of clinical symptoms of Salmonella Gallinarum were observed in the CaFe-GMP additive group, the group suffered no mortality from the infection.
Changes in feed intake rate in layers experimentally infected with Salmonella Gallinarum
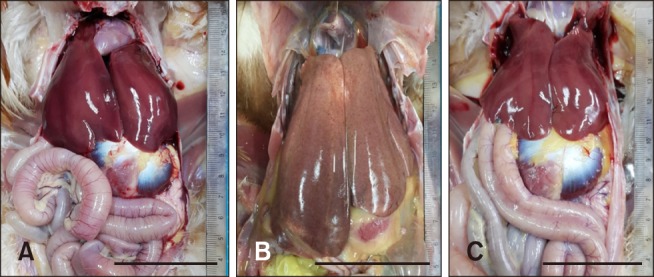
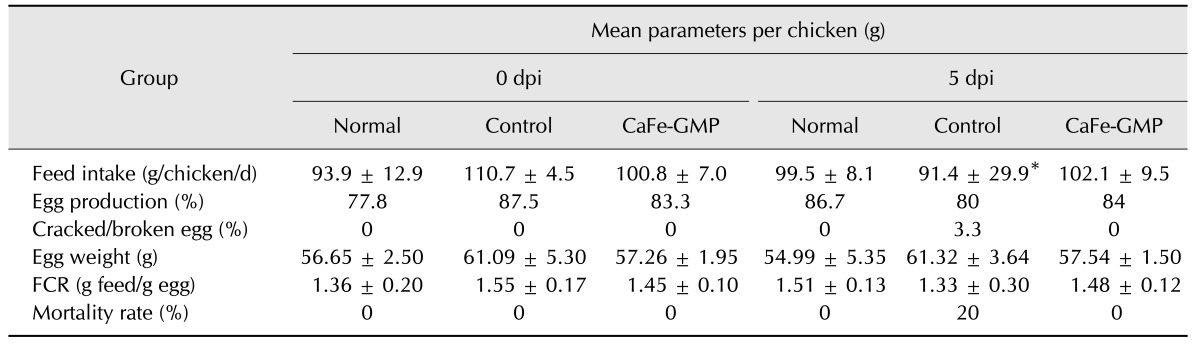
For each group, feed intake was compared at 0 dpi for all chickens and at 5 dpi for all surviving chickens. The mean feed group and (C) CaFe-GMP group. Scale bars = 5 cm. intake for chickens in the normal, CaFe-GMP additive, and control groups at 0 and 5 dpi are shown in Table 3. Compared to 0 dpi, feed intake in the control group decreased significantly by approximately 17.44% at 5 dpi, whereas in the CaFe-GMP additive group it increased slightly (1.2%) at 5 dpi; a non-significant change. The slight increase in feed intake of the CaFe-GMP additive group was less than that in the normal group, although neither increase was significant.
Evaluation of egg production and quality
Average egg production rate for the CaFe-GMP additive group was not significantly different from that of the control group before the Salmonella Gallinarum infection period (control group: 87.5%, CaFe-GMP group: 83.3%). After Salmonella Gallinarum infection, the CaFe-GMP additive group maintained a similar laying rate, whereas the laying rate for the control group decreased by 7.5% from that in the uninfected experimental period. The percentage of cracked or broken eggs for the control group after Salmonella Gallinarum infection was 3.3% at 5dpi, whereas no broken eggs occurred in the CaFe-GMP additive group during the experimental period (Table 3).
Macroscopic changes of spleen and liver
As shown in Table 4, the control group had significantly heavier spleens than those in the normal group. The CaFe-GMP additive group had a spleen weight significantly lower than that in the control group (p < 0.05); moreover, spleen size was similar to that in the normal group (data not shown). In addition, hepatomegaly was reduced in the CaFe-GMP additive group (Fig. 1). Furthermore, necropsy revealed that the CaFe-GMP additive group exhibited milder and less severe gross pathological abnormal changes when compared with those in the control group.
Bacterial clearance in internal organs
Surviving chickens from each group were sacrificed at 5 dpi. The liver, spleen, and feces from the cecum were aseptically collected and tested to determine whether Salmonella Gallinarum used for the challenge could be re-isolated from organs of the experimental chickens. The liver, spleen, and feces samples were negative for Salmonella Gallinarum in the normal group. In the control group, bacteria were re-isolated at 5 dpi from the liver, spleen, and cecum, indicating a systemic infection had occurred. As shown in Table 5, the numbers of Salmonella Gallinarum colony forming units in the CaFe-GMP additive group were considerably lower at 5 dpi in the liver (45.7%, p < 0.06), spleen (57.4%, p < 0.006), and feces from the cecum (96.6%, p < 0.057) than those in the control group. In particular, Salmonella Gallinarum detection was significantly cleared in excreted feces on sterilized paper litter from the housing units of the CaFe-GMP additive group (99.9%, p < 0.001%) at 5 dpi when compared with that of the control group.
Discussion
Avian systemic salmonellosis is primarily caused by the Salmonella Gallinarum strain and results in the development of fowl typhoid disease [7]. Fowl typhoid can be acute or chronic and usually results in a high mortality rate [2]. Symptoms include diarrhea, intestinal and womb bleeding, and enlargement of the liver and spleen.
Antibiotics may be used as feed supplements to improve physical performance by suppressing subclinical disease challenge in industrial animals [8]. However, there is worldwide concern regarding the overuse of antibiotics because of the development of multi-antibiotic resistant bacteria and resistance genes and their spread from animals to humans through consumption of food products that contain antibiotic residues [16]. Hence, there is an urgent need to identify natural-origin feed supplements that can reduce antibiotic use. Several studies have demonstrated that prevention of Salmonella colonization in chickens can be achieved by feeding with prebiotics, such as saccharides, organic acid, and inorganic minerals [142425283738].
Mineral additives in poultry feed are needed for bird survival and for efficient poultry production. The mineral requirements of laying hens are well reported [22], but information on trace mineral requirements and their related effects on various aspects such as immune function, resistance to disease, growth, and carcass composition is limited. In this study, we evaluated the effect of a CaFe-GMP feed supplement on the health and egg quality in layers experimentally infected with Salmonella Gallinarum.
Our results demonstrated that administering 0.16% CaFe-GMP as a feed additive had a beneficial effect on body weight in layers after Salmonella Gallinarum infection. It has been reported that mice on an iron-deficient diet gained less weight and ate less [32], suggesting that chickens fed a diet with an iron additive may exhibit less weight loss during intestinal inflammation by Salmonella Gallinarum infection.
Salmonella Gallinarum causes a high mortality rate in fowl [9]. However, in the present study, mortality did not occur in the Salmonella Gallinarum-infected CaFe-GMP additive group during the experimental period. Moreover, clinical signs of an infection were delayed, and the severity of related symptoms was lower in the CaFe-GMP additive group than in the control group. In the control group, there was severe, creamy whitish diarrhea observed from 4 dpi onwards, whereas in the CaFe-GMP additive group there were few typical clinical symptoms of Salmonella Gallinarum infection observed. In addition, the CaFe-GMP additive group had no fatalities. These results are similar to previous results showing that parenteral administration of complexed iron can enhance survival in chicks infected with Salmonella Gallinarum endotoxin (LPS) [30].
With regard to feed intake, the amount of feed consumed in the control group abruptly decreased, supporting results reported by Kim et al. [17]. In the Salmonella Gallinarum-infected CaFe-GMP additive group, there were no significant changes in feed consumption during the experimental period. Moreover, our results showed that the CaFe-GMP additive group displayed no significant change in growth performance.
Infection of fowl by Salmonella Gallinarum results in a decline in egg production rate and broken eggs [1131]. Interestingly, the infected CaFe-GMP additive group retained a normal egg production rate and broken eggs were not observed during the experimental period. Similarly, others have shown that dietary supplementation with a Fe-soy proteinate effectively improved egg quality [28], and dietary supplementation of an organic mineral mixture increased the bioavailability of minerals compared to that from inorganic sources [37]. CaFe-GMP supplementation may protect against the penetration of Salmonella spp. into the oviduct of layers and, therefore, could increase economic profits by maintaining egg production and hatchability in Salmonella-infected layers.
Salmonella infection in chickens occurs at all ages and is characterized by severe hepatomegaly and splenomegaly accompanied by anemia, septicemia, and a liver with a bronzing aspect [29]. Gross changes were observed in the organs of the control group, mainly in the liver, which was green-yellowish in color, larger, and more friable than that in normal layers. There was also spleen congestion and splenomegaly observed in the control group. In the CaFe-GMP additive group, all live chickens exhibited normal organ appearance.
We also examined whether Salmonella Gallinarum could be re-isolated, post-challenge, from organs of the experimental chickens. In the control group, bacteria were re-isolated at 5 dpi from the liver, spleen, and feces from the cecum, which indicated the occurrence of an infection. The numbers of Salmonella Gallinarum in all tested tissues of the CaFe-GMP additive group were significantly lower than those in the control group. Moreover, mortality in the CaFe-GMP additive group was delayed compared to that in the control group. Several reports have demonstrated that dietary iron intake lessens intestinal inflammatory responses and the pathology of enteric infection by foodborne bacterial pathogens, such as the Salmonella enterica serovar Typhimurium in mice [1834]. In addition, iron deficiency has been associated with an impaired immune response, but it can also substantially increase resistance against intracellular pathogens, probably due to an increase in nutritional immunity [35]. Because dietary iron status can affect the immune response, it is likely that the array of antimicrobial defenses secreted from the intestinal mucosa is also affected. Additionally, in the present study, necropsy revealed that the CaFe-GMP additive group had milder and less severe abnormal gross lesions and pathological changes than those observed in the control group, indicating that the CaFe-GMP feed supplement produced bacterial clearance as well as resistance against Salmonella Gallinarum.
Our findings suggest that adding 0.16% CaFe-GMP to the diet of layers can improve clinical symptoms of Salmonella Gallinarum, reduce diarrhea, and result in mild and muted abnormal gross pathological changes in liver and spleen. In addition, CaFe-GMP feed supplementation enabled bacterial clearance from internal organs and maintained egg laying production in layers experimentally infected with Salmonella Gallinarum. Consequently, a CaFe-GMP feed supplement may be useful in preventing salmonellosis in the poultry industry.


 XML Download
XML Download