

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Emergence and Evolution of HPAI H5Nx Clade 2.3.4.4
Wild birds, particularly those belonging to the orders Anseriformes (waterfowl, i.e., ducks, geese, and swans) and Charadriiformes (gulls and shorebirds), have long been recognized as natural hosts and reservoirs for all type A influenza viruses, also known as low pathogenicity avian influenza (LPAI) viruses [51]. These LPAI viruses have been isolated from more than 100 wild bird species belonging to more than 25 families, which confirms that LPAI viruses have a global distribution within free-living aquatic avian populations [8]. Highly pathogenic avian influenza (HPAI) viruses are not normally present in wild bird host reservoirs [38]. The first cases of HPAI, previously termed fowl plague, were reported in 1878 in chickens in northern Italy, which were followed by geographically dispersed outbreaks through the late 1800s to the 1950s in Europe, Asia, Africa, and North and South America [45]. In general, since 1959, HPAI outbreaks have been more geographically restricted with 42 distinct HPAI epizootics. The HPAI viruses arose from H5 or H7 LPAI viruses [46] after circulation for weeks to years in gallinaceous poultry and have developed specific mutations in the hemagglutinin (HA) gene that have conferred the phenotypic trait of high pathogenicity [47]. Most previous HPAI virus epizootics have been geographically limited, involving farm-to-farm spread, and were eradicated from poultry by stamping-out programs; i.e., the HPAI virus did not circulate in wild birds. However, an H5N1 HPAI virus was detected in 1996 in a domestic goose in Guangdong China (Gs/GD lineage) and, to date, unlike previous HPAI outbreaks, this lineage caused deaths in wild birds, poultry, and humans and has spread to over 80 countries in Asia, Europe, Africa, and North America, drastically changing the perspective on HPAI biology. The severity, size, and broad geographic distribution of the Gs/GD epizootic merits the term panzootic [45].
The H5N1 Gs/GD lineage HPAI virus has evolved into 10 genetically distinct virus clades (0–9) and multiple subclades [52]. Since 2008, HPAI subtypes H5N2, H5N5, and H5N8 bearing the genetic backbone of the Gs/GD lineage H5 clade 2.3.4 have been identified in poultry in China, especially in domestic ducks and poultry in live poultry markets, and those subtypes have subsequently evolved into different subclades including 2.3.4.4 [125355]. The H5 clade 2.3.4.4 HPAI viruses have undergone genetic reassortment with other clades of H5N1 viruses and with other local LPAI virus subtypes. In early 2014, outbreaks of novel reassortant H5N6 and H5N8 of clade 2.3.4.4 were reported in poultry and wild birds in China, Laos, and Vietnam [48] and in Japan and Korea [28], respectively. Previous studies have confirmed the wide geographic dispersion of Gs/GD lineage H5 clade 2.3.4.4 HPAI viruses since late 2014 and the movement of this virus from East Asia to North America, West Asia, and Europe [1026]. Genetic analyses have recently indicated that, since late 2012, the H5 clade 2.3.4.4 HPAI viruses have evolved into four distinct genetic groups [22]. Group A comprises H5N8 viruses identified in China in early 2014 and Korea, Japan, Taiwan, Canada, the United States (US), and European countries in 2014–2015. Group B comprises H5N8 viruses identified in China in 2013–2014 and Korea in 2014. Group C comprises H5N6 viruses identified in China and Laos in 2013–2014, and H5N1 viruses identified in China and Vietnam in 2014. Group D comprises H5N6 viruses identified in China and Vietnam in 2013–2014.
Spread of H5N8 HPAI Clade 2.3.4.4 Group A, 2014–2015
The Korean H5N8 outbreak in January 2014 was the first H5N8 epizootic reported outside China [28]. Migratory birds were suspected to have a key role in the introduction of groups A and B from Eastern China and in the subsequent spread during the 2014 Korean outbreak [14]. During the initial stage of this outbreak, identical viruses from each group were identified in poultry farms and wild birds near Donglim Lake, a wintering site for migratory birds in Korea, thereby suggesting that two different H5N8 HPAI viruses were introduced into Korea by wild birds during the same period [32]. As time passed, outbreaks caused by the Group A H5N8 HPAI virus predominated in the western region of Korea. Subsequently, the Group A H5N8 virus was identified in a long-distance migrant (Eurasian wigeon, Anas penelope) in Eastern Siberia in September 2014 [31] and in multiple wild bird species in East Asia, Europe, and the west coast of North America in November and December 2014 (Fig. 1) [1026]. Results of phylogenetic studies indicated that Group A viruses further evolved into three distinct subgroups in circumpolar regions and disseminated to various regions: icA1 (Europe/East Asia), icA2 (North America/East Asia), and icA3 (East Asia) subgroups. In Korea, Group A H5N8 viruses induced continuous outbreaks for 28 months, from January 2014 to April 2016, and mainly affected domestic duck farms [1340]. Three subgroups of H5N8 viruses (icA1–3) were re-introduced to Korea by migratory birds, and another subgroup of Group A H5N8 viruses was identified in Korea; a subgroup that had been maintained in poultry farms in Korea during 2014–2016 [1921]. After dissemination from Korea to various regions, these Group A H5N8 HPAI viruses generated a variety of reassortant viruses that shuffled gene segments with those of prevailing LPAI viruses from wild aquatic birds in North America and Taiwan [2227]. In North America, the Gs/GD lineage Group A H5N8 virus underwent reassortment with North American LPAI viruses to produce H5N1, H5N2, and H5N8 reassortant HPAI viruses [22]. The reassortant H5N2 HPAI virus predominated in the 2014–2015 US outbreak, mainly affecting commercial turkey and layer farms. This strain was eradicated from US poultry in June 2015 and was rarely detected in wild birds in 2016 [25]. In Taiwan, H5N2, H5N3, and H5N8 subtypes bearing the HA segment of the H5 clade 2.3.4.4 Group A have caused outbreaks in poultry and wild birds since January 2015 [27].
Spread of H5N8 HPAI Clade 2.3.4.4 Group B, 2014–2017
The Group B H5 clade 2.3.4.4 HPAI viruses have been detected in China since 2013 and are phylogenetically distinct from Group A H5N8 viruses that spread globally during 2014–2015 [752]. In early 2014, outbreaks of H5N8 clade 2.3.4.4 groups A and B were reported in Korea, followed by outbreaks of Group A H5N8 in East Asia, Europe, and North America in late 2014 (Fig. 1) [2628]. Interestingly, there have been no further reports of Group B virus in Korea since the initial report in January 2014 [26]. In China, several Group B viruses reassorted with LPAI viruses and were reported during 2013–2014 mostly from domestic ducks and wild aquatic birds [729415356]. Novel reassortant Group B H5N8 viruses containing 5 Eurasian LPAI segments (PB2, PB1, PA, NP, and M) were identified from wild birds found dead in Qinghai Lake, China during May 2016 [30] and at Uvs-Nuur Lake in Siberia during June 2016 [24]. Subsequently, this novel reassortant Group B H5N8 strain has further reassorted with Eurasian LPAI viruses and disseminated from Siberia to be detected over a broad geographic region that includes Europe [36], Africa [39], the Middle East [9], and Asia [1832]. That dissemination may be the result of the southward autumn migration of waterfowl during 2016 and was similar in the spread of H5N1 clade 2.2 in 2005–2006 [35] and H5N8 clade 2.3.4.4 Group A in 2014 [2631] from Siberia to various countries in Eurasia.
Spread of H5N6 HPAI Clade 2.3.4.4 Group C, 2014 –2017
While Group A and B H5N8 viruses caused global outbreaks during 2014–2015 and 2016–2017, the Group C H5N6 viruses have caused outbreaks in China since 2013 [637], and Group C viruses have been isolated from various host species including wild birds, poultry, pigeons, pigs, cats, and humans [4]. The Group C H5N6 viruses disseminated to Laos and Vietnam in 2014 and Hong Kong in 2015 [54]. Recently, Bi et al. [4] reported that at least 34 distinct genotypes of H5N6 HPAI virus have been identified through surveillance of live poultry markets in China during 2014–2016. In autumn 2016, Group C H5N6 viruses belonging to the G1.1.9 genotype were identified in Korea [15,20,25] and Japan [34], followed by continuous outbreaks in domestic poultry and wild birds in Korea (Fig. 1) [4]. Multiple novel reassortant H5N6 viruses containing different PA and NS genes of Eurasian LPAI viruses have been reported from Korea, and those novel viruses seem to have likely originated from the H5N6 viruses circulating in Guangdong province of China [25].
Pathogenicity of H5Nx HPAI Clade 2.3.4.4 in birds
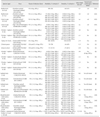
Numerous experimental infections have been performed to investigate the pathogenicity of clade 2.3.4.4 HPAI viruses in different bird species. To date, the majority of characterized viruses are Asian (Table 1) and North American (Table 2), but there are a few studies describing the pathobiology of natural cases of H5N8 HPAI virus infection in Europe.
Asian H5 HPAI clade 2.3.4.4
Subtypes H5N2, H5N6, and H5N8 clade 2.3.4.4 viruses isolated from apparently healthy ducks and geese in China were characterized in 4-week-old domestic Pekin ducks (Anas platyrhynchos domesticus) and layer chickens [44]. All reassortants caused systematic infection in ducks when challenged at 6 log10 mean egg infectious dose (EID50) and could be efficiently contact-transmitted with high titers shed consistently through oropharynx and cloaca. The infected ducks showed severe neurologic signs. Severe hemorrhages and necrosis in the heart and pancreas were observed at necropsy. Acute necrosis of cardiac muscle cell, neuronophagia, and multifocal necrosis of acinar cells were present, with viral antigen detected in alveolar epithelial cells, cerebrum, brain stem, and pancreas. Interestingly, virus titers in the brains of H5N6- and H5N8- inoculated ducks were significantly higher than those of H5N2- infected ducks. Similar results were obtained in chickens [44]. However, in accordance with other studies, Sun et al. [44] concluded that these H5 reassortants have reduced virulence compared to the parental H5N1 virus as evident by a decrease in the intravenous pathogenicity index (IVPI) and a lengthened mean death time. In another study, three H5N8 viruses isolated from apparently healthy domestic geese at a wholesale live-bird market in Eastern China in 2014 were demonstrated to be highly virulent to 4-week-old chickens and mallard ducks (Anas platyrhynchos) when challenged at 6 log10 EID50 [29]. All challenged birds were capable of efficiently transmitting to contacts, and showed systemic infection with myocardial necrosis as the main lesion [29].
Tanikawa et al. [49] compared the pathogenicity of chicken (Miyazaki/2014) and wild duck (Chiba/2014) Japanese Group A H5N8 viruses in chickens. All chickens inoculated with 4–6 log10 EID50 of Chiba/2014 became infected and died by 13 days post-challenge, whereas 25% and 75% of the chickens inoculated with 4 or 5 log10 EID50, respectively, of Miyazaki/2014 became infected and died. Both isolates replicated to high titers in both trachea and cloaca [49]. A Group A H5N8 virus isolated from a Japanese chicken farm during an outbreak in April 2014 was tested in chickens and 10-week-old Pekin ducks [16]. The challenge was lethal only in chickens inoculated with the highest dose tested (6 log10 EID50), and the virus was systemically present in tissues. In contrast, all doses (2, 4, 6 log10 EID50) resulted in asymptomatic infections in Pekin ducks, with consistent oral and cloacal shedding [16]. Recently, the pathogenicity of two Group A H5N8 viruses isolated from wild migratory birds in late 2014 in Japan was examined [50]. The viruses possessed atypical HA cleavage sequences: a swan isolate, Tottori/C6, had a novel polybasic cleavage sequence PLGERRRKR (the polybasic HA cleavage site is the prime virulence determinant of avian influenza viruses) and a mandarin duck (Aix galericulata) isolate, Gifu/01, had a heterogeneous mixture of consensus PLRERRRKR and variant PLRERRRRKR sequences. The Gifu/01 isolate caused higher and earlier mortality, higher viral titers systemically in tissues, and more severe lesions than those in the Tottori/C6 isolate after intravenous inoculation in chickens (6 log10 EID50), suggesting that the novel cleavage motif of the Tottori/C6 isolate did not directly affect the infectivity of the virus [50].
The pathogenicity of the Group A H5N8 virus that caused outbreaks in wild birds in Korea in 2014 was examined in a variety of bird species [23]. Although all species were permissive for infection and virus shedding at a challenge dose of 6 log10 EID50, severe infection was observed in layer chickens and Japanese quail (Coturnix japonica), mild infection in Korean native chickens, and asymptomatic infection in Muscovy ducks (Cairnia moschata domestica). Transmission to contact-exposed birds occurred in all tested species but was sporadic in chickens, intermediate in quail and Korean chickens, and consistent in 8-week-old Muscovy ducks [23]. When compared to the previously reported H5N1 HPAI viruses, Lee et al. [23] concluded that the Group A H5N8 virus has low pathogenicity and transmissibility in poultry species. Recently, a Group A H5N8 isolate from a broiler duck farm showed efficient respiratory replication in adult mandarin ducks and transmission to contacts at a challenge dose of 6.5 log10 EID50, along with asymptomatic infection and gross lesions only in a few tissues [17].
North American H5 HPAI clade 2.3.4.4
Representative Eurasian Group A H5N8 and its reassortant Group A H5N2 viruses detected in wild birds at the end of 2014 in North America have been tested in a variety of avian species [3,5,42,43]. Recently, the pathogenesis of these clade 2.3.4.4 wild bird viruses was investigated in juvenile and adult ruddy ducks (Oxyura jamaicensis) and juvenile lesser scaups (Aythya affinis), as representatives of wild diving duck species [43]. These species showed asymptomatic infection at relatively low challenge doses (as low as 2 log10 EID50 for adult ruddy ducks and 4 log10 EID50 for juvenile ruddy ducks and lesser scaups), with oral and cloacal virus shedding and systemic virus presence in infected tissues. Age differences in susceptibility to disease were also observed [43]. Interestingly, shorter shed duration and lower titers would suggest that these diving ducks are less susceptible to efficiently transmit the viruses than mallards [5]. In another study using these same North American wild bird viruses in chickens, pathobiological features were consistent with HPAI virus infection, but other results such as susceptibility to infection only at high challenge doses (4 or 6 log10 EID50) and lack of contact-exposed transmission indicated that these wild bird viruses were poorly adapted to chickens [3]. Interestingly, the novel reassortant H5N1 virus that was isolated from backyard chickens in British Columbia, Canada at the end of 2014 was shown experimentally to have a wide host range [1]. This virus was highly pathogenic to chickens, as determined by the IVPI value [1]. In turkeys (Meleagris gallopavo), juvenile Muscovy ducks, and adult Chinese geese (Anser cynoides domesticus), the virus caused systemic infection at challenge doses of 3–4 log10 plaque forming units and was efficiently transmitted to and caused mortality in naïve contact birds [1].
Subsequently in 2015, the H5N2 HPAI virus outbreak in the US Midwest affected commercial turkey and layer farms. Experimentally, four poultry H5N2 viruses exhibited higher virus adaptation to chickens than that of the wild duck 2014 US index H5N2 virus, but they still transmitted poorly to contact-exposed chickens [5]. At the same time, these poultry isolates retained high adaptation to mallards; two of the H5N2 poultry isolates had similarly high infectivity and transmissibility as those of the index H5N2 virus, with mortality (1 of 8 at a challenge dose of 6 log10 EID50) occurring with only one of two poultry H5N2 isolates. Virus antigens were present in parenchymal cells of many organs including cardiac myocytes, hepatocytes, microglial cells and neurons, lung, and kidney tubular epithelial cells. Viral antigen staining in capillary endothelial cells was uncommon and was restricted mainly to capillaries in eyelid and air capillaries of lung [5]. Experimentally, both early (2014) and late (2015) occurrences of H5N2 viruses in turkeys had unusually long mean death times, high levels of virus in feces, and increased adaptation of the later viruses, which collectively suggest that this species was permissive for infection and may have contributed to the rapid spread of the virus during the peak of the outbreak [42].
To assess potential genetic resistance of broilers and/or age-related effects that could explain the lack of affected broiler farms in the 2015 US outbreak, the pathogenesis and transmission of one of the poultry H5N2 isolates were investigated in commercial broilers of different ages [2]. In that study, chickens died, shed high virus titers, and enabled transmission only following the highest dose challenge (6 log10 EID50), but overall age was not a determinant factor in susceptibility. Taking into account all of the studies reported to date, it seems that the broiler breed is less susceptible to Midwestern US H5N2 viruses than the layer breed but similarly susceptible as turkeys. Therefore, genetic resistance of broilers to infection may account only partially for the lack of affected broiler farms in the US Midwest, with other contributing factors including fewer outside to on-farm exposure to contacts, type of production management system, and enhanced biosecurity [2].
European H5 HPAI clade 2.3.4.4
To date, there are no published reports on experimental studies of the HPAI clade 2.3.4.4 viruses that have affected European domestic and wild birds since 2014. However, two natural infections with extensive pathobiological characterization have been reported. An outbreak of Group A H5N8 virus was detected in a breeder duck farm in the United Kingdom in November 2014 [33]. The holding, with approximately 6,000 breeder ducks aged approximately 60 weeks, showed a gradual reduction in egg production and increased mortality over a seven-day period. Post-mortem examination revealed frequent fibrinous peritonitis, severely hemorrhagic ovarian follicles, occasional splenic and pancreatic necrosis, and a high incidence of mycotic granulomas in air sacs and lung. Low-to-moderate levels of virus were detected mainly in respiratory and digestive tracts, with minor involvement of other organs. Although histopathological examination confirmed the gross pathology results, intralesional viral antigens were not detected by immunohistochemical analysis. Immunolabeled cells were rarely and only present in inflamed air sacs and serosa, usually superficial to granulomatous inflammation. Abundant bacterial microcolonies were observed in hemorrhagic ovaries and oviduct. The authors suggested that the limited viral tissue distribution and the presence of intercurrent fungal and bacterial infections may indicate a minor role for H5N8 virus in clinical disease in layer ducks [33].
In Germany, a Group A H5N8 virus not only affected wild birds and commercial poultry but also birds kept in a zoo in northeastern Germany [11]. Approximately, 500 birds of 94 different species were kept in the Rostock Zoo, Germany in the autumn of 2014. Only a few individual white storks (Ciconia ciconia) showed clinical signs and eventually died in the course of the virus infection, although subsequent in-depth diagnostic investigations showed that other birds kept in the same compound as the white storks were acutely positive for or had undergone subclinical infection with the H5N8 virus. The most striking lesions in the three white storks found dead were necrosis in liver, kidneys, spleen, and pancreas. Only one of the 7 scarlet ibis (Eudocimus ruber) that were investigated tested positive for H5N8 viral RNA and showed liver and adrenal necrosis [11].
Concluding Remarks
Since 2014, HPAI clade 2.3.4.4 viruses have spread rapidly and globally through wild birds and have evolved through reassortment with prevailing local LPAI viruses. A wide range of avian species, including wild and domestic waterfowl, domestic poultry, and even zoo birds, appear to be permissive for infection by and/or transmission of clade 2.3.4.4 viruses. In general, clade 2.3.4.4-infected birds exhibit clinical disease, mortality, and pathological features that are indicative of a typical HPAI virus infection, although several studies have pointed out that these H5 reassortants have reduced virulence compared to the parental Gs/GD H5N1 virus. Interestingly, clade 2.3.4.4-infected waterfowl may either succumb to infection or act as asymptomatic spreaders, even with viruses that are optimally adapted to domestic poultry. The wide host range of clade 2.3.4.4 viruses, which can enable unrecognized virus spread, might explain the successful recent globalization of this lineage. The emergence of novel reassortant viruses and their circulation among bird populations are of great concern because of the potential for virus dissemination by intercontinental wild bird migration. Therefore, in places where HPAI has become endemic in domestic poultry, explicit efforts should be taken to eradicate HPAI by using a combination of preventive and control measures that include stamping out, appropriate vaccination with post-vaccination monitoring, movement control, and enhanced biosecurity. In addition, intensified surveillance and further characterization of HPAI viruses can provide insight into preventing future outbreaks and assessing zoonotic potential.


 XML Download
XML Download