

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Currently, three subtypes (H1N1, H3N2 and H1N2) of swine influenza virus (SwIVs) are circulating in swine populations throughout the world. The 1968 Hong Kong pandemic was caused by an outbreak of the H3N2 strain, which has been transmitted to domestic pigs and undergone frequent reassortment [17]. However, H3N2 SwIV infection was reported sporadically in the swine population until a swine-adapted reassortant virus emerged in 1998, which rapidly spread through the US swine population and caused severe respiratory and reproductive diseases [19]. Most recently, a novel reassortant H3N2 SwIV variant with the M gene from the 2009 pandemic virus was isolated in humans in multiple US States [2]. Currently, influenza vaccine strains for humans are selected annually based on global epidemiological surveillance data. However, since there is no comprehensive surveillance of SwIVs, development of universal vaccines with broad cross-reactivity against SwIVs is urgently needed.
The extracellular domain of influenza M2 protein (M2e) is highly conserved among human influenza A viruses and considered an appropriate target for the development of influenza vaccine with broad-spectrum protection [4]. Vaccination with M2e could protect animals against different subtypes of influenza viruses, including avian H5N1 influenza virus [18]. The protection was mediated by M2e-specific antibodies via antibody dependent cytotoxicity [9]. In addition, M2e monoclonal antibody could reduce the plaque size of influenza A virus and inhibit growth of the virus in vitro [81624]. However, the human body could not induce high levels of antibody against M2e due to the poor immunogenicity of M2e. Therefore, development of a conjugate vaccine that could increase the production of antibodies against M2e in the swine population is a promising way to develop M2e based universal influenza vaccine.
Surface protein hemagglutinin (HA) and neuraminidase (NA) are the major antigens of influenza virus, and the production of antibodies against HA is crucial for immune protection [5]; therefore, we selected the HA and NA of a prevailing H3N2 isolate and six internal genes from A/Puerto Rico/8/34 (PR8) to generate a high-growth reassortant H3N2 influenza seed virus (rgH3N2) [20]. We designed a novel tetra-branched MAP based vaccine, which was constructed by fusing four copies of M2e to one copy of foreign T helper (Th) cell epitope. In this study, mice were vaccinated with rgH3N2 inactivated vaccine combined with the novel M2e-MAPs to determine whether supplementation with M2e-MAP improved the immune response and provided cross-protection against swine and human viruses.
Materials and Methods
Mice
Six-week-old female BALB/c mice were purchased from SLAC Laboratory Animal (China) and housed in a specific pathogen-free (SPF) facility following the approved animal care protocols. The mice were acclimated to their new environment for one week prior to immunization. All experimental protocols involving mice were approved by the Chinese Ministry of Agriculture and the Review Board of Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences (permit No. SYXK 2011–0116).
Viruses and peptide
A/Swine/Guangdong/164/06 (SwGD164; H3N2), A/Swine/Heilongjiang/1/05 (SwHLJ1; H3N2) and A/Swine/Guangdong/96/06 (SwGD96; H1N1) were isolated from pig farms in China [22]. PR8 (H1N1) is a mouse-adapted virus. All viruses were grown for 48 hours at 37℃ in the allantoic cavities of 10-day-old SPF chicken embryo eggs. The novel tetra-branched M2e-MAP was constructed by fusing four copies of a universal sequence, which is an extracellular part of the M2 protein, with one copy of promiscuous Th cell epitope by a lysine tree with two additional Lys residues defining a putative cleavage site for cathepsin D [12]. It was synthesized by GenScript (China). M2e-MAP was prepared at 10 mg/mL in distilled water and stored at −20℃. Its construction and sequence are shown in Fig. 1.
Generation of a high growth reassortant virus (rgH3N2)
rgH3N2 harboring the HA and NA genes from SwGD164 and six internal genes from PR8 was generated by plasmid-based reverse genetics as previously described [20]. Briefly, one microgram of each plasmid was added into 250 µL Opti-MEM (Invitrogen, USA) and vortexed. Next, 16 µL of the transfection reagent Lipofectamine 2000 (Invitrogen) were added into 250 µL Opti-MEM and mixed gently. Five minutes later, the diluted transfection reagent was mixed with the diluted plasmids. The DNA-transfection reagent mixture was kept at room temperature for 20 min, then added directly to a monolayer of 293T cells in a 6-well plate (Costar; Corning, USA). After 6 h of incubation at 37℃ in 5% CO2, the medium was replaced with 2 mL fresh Opti-MEM, and the plate was incubated as described above for 48 h. The supernatant was subsequently inoculated into the allantoic cavity of 10-day-old embryonated SPF eggs. The allantoic fluid was harvested after 48 h of incubation at 37℃, after which the virus was identified by hemagglutination assay. Reverse transcription PCR (RT-PCR) and re-sequencing confirmed that the genome of the rescued virus was identical in sequence to the cDNA in the plasmids used for its rescue [20]. The rescued viruses were then confirmed by sequencing. The titer of the rgH3N2 virus was determined in 10-day-old SPF embryonated chicken eggs, and the 50% embryo infective dose (EID50) was calculated by the Reed-Muench method.
Vaccine preparation
The rgH3N2 virus was propagated in the allantoic cavity of 10-day old embryonated SPF chicken eggs and inactivated with 0.1% (v/v) formalin for 48 h at 4℃. The virus was then purified by sucrose density gradient centrifugation [11].
The novel M2e-MAP was synthesized and stored at −20℃. One dose of M2e-MAP combined vaccine containing 10 µg of each inactivated rgH3N2 and M2e-MAP (in 50 µL sterile PBS) was mixed and emulsified with the same volume of Freund's incomplete adjuvant (FIA; sigma, USA) or Freund's complete adjuvant (FCA; sigma).
Immunization and virus challenge
A total of 120 female BALB/c mice were randomly divided into four groups (n = 30 in each group), immunized subcutaneously with immunogen plus FCA and boosted with the same amount of immunogen in FIA at 2-week intervals. Mice in group 1 (M2e-MAP+rgH3N2+Freund) received one dose of M2e-MAP combined vaccine. Mice in group 2 (rgH3N2+Freund) were immunized with one dose of the vaccine as prepared above. Mice in group 3 (rgH3N2) and group 4 (Freund) were immunized with inactivated virus and Freund alone as controls, respectively. The immunization schedule of mice in the experiment is shown in Table 1. Serum samples from five mice in each group were collected at 0, 1, 2, 3 and 4 weeks after the first immunization. Mice (n = 10 for each virus) were anesthetized and challenged intranasally with 106 EID50 of SwGD96 and SwHLJ1, as well as 10 LD50 of PR8 on day 14 after the booster. The loss of body weight and symptoms were monitored daily and the survival curves were analyzed by the Kaplan-Meier method.
Antibody detection
Enzyme-linked immunosorbent assay (ELISA) was applied to determine titers of antibody specific to rgH3N2 and M2e-MAP in serum samples collected on days 7, 14, 21 and 28 post primary immunization as described previously [21]. Hemagglutination inhibition (HI) assay was performed according to the WHO Manual on Animal Influenza Diagnosis and Surveillance (World Health Organization, Geneva, Switzerland). Neutralization tests (NTs) were performed according to Kida et al. [10].
Virus titers in lung tissues
Lung tissues from euthanized mice were removed at 3 days post infection (dpi) and homogenized in 500 µL of ice cold PBS. Virus titers in mice lungs were determined by EID50 titration and quantitative real-time PCR as previously described [20]. The specific primers and labeled fluorescent-probe were as follows: InfA-forward: 5-GACCGATCCTGTCACCTCTGAC-3; InfA-rerverse: 5-AGGGCATTCTGGACAAAGCGTCTA-3; TaqMan probe: 5-FAM-TGCAGTCCTCGCTCACTGGGCACGBHQ-3.
Histopathological analysis
The lung tissues of challenged mice were collected on day 4 and fixed in 10% neutral buffered formalin, and histopathological changes of the mouse lungs were examined by hematoxylineosin stain and observed under a light microscope.
Statistical analysis
Data were analyzed and charted with Excel and Graphpad Prism 5. The survival curves were analyzed by the Kaplan-Meier methods, and the significance of differences in overall survival between groups was determined by the log-rank test. Statistical significance was determined by two-way ANOVA.
Results
Characterization of the reassortant virus rgH3N2
The rgH3N2 virus harboring the HA and NA genes from SwGD164 and six internal genes from PR8 was generated by plasmid-based reverse genetics and confirmed by sequencing analysis. The HA titer of rgH3N2 reached 512 HAU/50 µL, which was 2 fold higher than that of SwGD164. The rgH3N2 virus (106.7EID50/mL) grows to a higher titer than SwGD164 in embryonated chicken eggs.
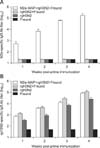
M2e-MAP combined vaccine induced strong antibody responses
M2e-MAP combined vaccine induced strong M2e-specific and rgH3N2-specific antibody responses in mice (Fig. 2). The titer of M2e-sepcific antibody reached 1 : 103 at 7 days post primary immunization and increased to the highest titer of over 1 : 105 at 14 days post boost. In contrast, only background levels of antibody were detected in mice vaccinated with Freund's alone (Fig. 2). Both the antibody titers to rgH3N2 and M2e-MAP increased after the boost. The antibody titer of rgH3N2 in group 3 was lower than the one emulsified with Freund (groups 1 and 2) at each sampling point.
The HI and NT tests were performed on serum collected 2 and 4 weeks post primary immunization. As shown in Table 2, HI antibodies and neutralizing antibodies were detected in all groups except for the control. The HI and NT titers were almost equal in groups 1 and 2, where the average titers of HI and NT were found to be 256 and 103 after the first immunization, respectively. The mean titers of HI and neutralizing antibodies were up to 1,152 and 104 until viral infection, respectively. In group 3, the average titer of HI antibodies was 160 and that of the neutralizing antibody was 102 after the first immunization. Until viral infection, the mean titers of HI and neutralizing antibody were up to 512 and 103, respectively. The above results indicated that immunization with M2e-MAP combined with H3N2 inactivated vaccine induced high levels of humoral responses in mice.
M2e-MAP vaccination limits viral replication
Mouse lungs were collected on 3 dpi and the virus titers were determined by EID50 titration and quantitative real-time PCR (Fig. 3). No detectable influenza virus was present in the lungs of mice challenged with SwHLJ1 (H3N2) or SwGD96 (H1N1) (< 1.0 log10 EID50/mL), and the titer decreased to 103.25 ± 0.35 EID50/mL in PR8 challenged mice (Table 2). The addition of M2e-MAP significantly increased the ability of the vaccine to reduce heterologous virus replication in group 1. Real-time quantitative PCR indicated M2e-MAP combined vaccine provides strong protection against PR8 (H1N1), SwGD96 (H1N1) and SwHLJ1 (H3N2) with significant differences in viral titers among four groups (p < 0.001).
M2e-MAP vaccination reduces lung lesions
Mouse lungs were collected for histopathologic examination at 3 dpi. In groups 2, 3 and 4, moderate to serve broncho-interstitial pneumonia was observed (Fig. 4). In groups of mice challenged with PR8, lung tissues were damaged and inflammatory cell infiltration was observed in large areas in the control group (panel J in Fig. 4). However, the lung tissues of mice that were challenged with PR8 in group 1 appeared relatively normal aside from slight alveolar dilatation (panel A in Fig. 4). No histopathologic changes were observed in the group 1 mice challenged with SwGD96 (panel B in Fig. 4) or SwHLJ1 (panel C in Fig. 4).
Body weight and survival curve
The body weight and survival rate of the mice that received lethal challenge (10LD50) of PR8 were monitored for 14 days post challenge (Fig. 5). Survival rates of mice in groups 1, 2 and 3 were 100%, 40% and 30%, respectively (p = 0.0002) (Fig. 5A). All mice in group 4 died on day 9. As shown panel B in Fig. 5, the body weight of mice in group 1 decreased slightly at first, then increased on day 10. Lethargy, loss of appetite, fever and shiver were observed in groups 2, 3 and 4 at 2 dpi.
Discussion
Influenza A viruses infect a broad range of species, with avian and swine strains presenting the greatest potential for interspecies transmission. Since the 1990s, H3N2 triple-reassortant viruses with genes derived from human (HA, NA, and PB1), swine (N, M, and NS), and avian (PB2 and PA) influenza viruses have caused outbreaks of respiratory disease in pigs throughout North America [1425]. Yu et al. [22] summarized and reported the co-existence of wholly human-like H3N2 viruses, double-reassortant H3N2 viruses, and triple-reassortant H3N2 viruses in pigs in China for the first time by analyzing the homology among eight genes of swine influenza A (H3N2) viruses found in China from 1970 to 2006. The HA and NA genes in SwGD164 used in this study showed high homology with the prevailing H3N2 SwIV, but its fecundity in chicken eggs was low. To improve the replication ability of SwGD164 in chicken eggs, we generated a high-growth rgH3N2 virus that contains the HA and NA from SwGD164 and six internal genes from the PR8 virus by plasmid based reverse genetics. The HA titer of rgH3N2 was significantly improved compared to SwGD164.
It has been reported that cross-protective immunity might play an important role in protection against emerging influenza pandemics [6]. The specific conserved epitopes of cytotoxic T lymphocytes play a role in heterologous protection [13]. The vaccination of seasonal influenza vaccine (H3N2/H1N1) provided cross protection to heterologous virus (H5N1), and CD4T cells were involved [7]. The antibody to conserved nucleoprotein can also provide heterologous protection [1]. The highly conserved M2 protein of influenza virus is an ideal target of universal vaccine with cross protection; however, influenza virus infection or immunization cannot induce specific antibody to M2 protein [918], which could not be largely synthesized in infected cells and were occasionally packaged into nucleocapsids. It has been reported that anti-M2e IgG can eliminate infected cells by antibody-dependent cell mediated cytotoxicity or complement-mediated phagocytosis, in which the natural killer cells, alveolar macrophages and dendritic cells play an important role [39]. Thus, M2e-based vaccines were developed and have been reported to provide broad cross protection against divergent or heterologous influenza viruses [121524].
In the present study, we developed a novel tetra-branched M2e-MAP and combined it with a high rgH3N2 inactivated vaccine. The mice that received the M2e-MAP combined vaccine were then infected with 106 EID50 of SwGD96 and SwHLJ1 and 10LD50 of PR8 through nasal inoculation. High titer of HI and neutralizing antibody induced by the vaccine provided effective suppression of replication of homologous SwHLJ1 virus (H3N2) and pathological changes in the lung, with a protection rate of 100%. Reassortant H3N2 vaccine also provided cross protection, determined by the internal gene, against heterologous virus PR8 (H1N1) (40%). These results suggest that the addition of M2e-MAP did not affect the generation of rgH3N2 specific HI and neutralizing antibodies. Moreover, vaccination of M2e-MAP combined vaccine reduced virus replication and pathological changes in the lungs, which further indicated that M2e-MAP may improve immune protection of vaccine against heterologous virus effectively. High titer of HI and neutralizing antibody induced by rgH3N2 + M2e can provide effective suppression of the replication of homologous SwHLJ1 virus (H3N2) and pathological changes in the lung, with a protection rate of 100%. In addition, reassortant H3N2 vaccine produced some cross protection, determined by internal gene, against heterologous virus PR8 (H1N1) (40%).
Taken together, the approach of developing M2e-MAP combined vaccines with cross-protection is feasible. Our study provided solid evidence that M2e-MAP can significantly enhance the protective ability of traditional inactivated vaccine against heterologous viruses. The M2e-MAP combined high growth reassortant H3N2 inactivated vaccine developed in this study could provide cross-strain protection and thus serve as a candidate for SI vaccine in China.






 XML Download
XML Download