

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Mycobacterial species other than those belonging to the Mycobacterium (M.) tuberculosis complex or M. leprae are known as nontuberculous mycobacteria (NTM), environmental mycobacteria, or atypical mycobacteria [14]. NTM are ubiquitous in the environment and induce opportunistic infections that can produce a wide range of diseases, and a large number of NTM have been isolated from mammals [20]. As opportunists, NTM cause acute or chronic infections in patients with cystic fibrosis [1], cancer [4], HIV [21] and immunosuppression [18]. In humans, NTM comprise an important clinical problem associated with significant morbidity, and their occurrence is increasing worldwide [35]. Accordingly, researchers are searching for an immunogenetic factor that will illuminate susceptibility to non-tuberculous mycobacterial disease. To identify new therapeutic targets and enable rational vaccine design, increased understanding of the molecular and cellular host defense mechanisms providing protective immunity against NTM is needed [17]. However, most studies of NTM conducted to date have focused on epidemiological investigations [35], drug sensitivity [11] and laboratory diagnosis [33]. Indeed, there have been few investigations of host immune mechanisms following infection or the reasons for which NTM can persist in the host. Therefore, there is an urgent need to clarify the infection mechanisms of NTM.
Innate immune responses play an important role in bacterial destruction; however, the chronic nature and high incidence of M. tuberculosis with low levels of CD4+ T cells indicates the importance of adaptive immune effectors [1729]. T helper 1 (Th1) cell-mediated immune responses can effectively remove Mycobacterium [26]. Following infection, a central control step against mycobacteria is activation of CD4+ T helper 1 cells, which produce effector cytokines such as tumor necrosis factor alpha (TNF-α) and gamma-interferon (IFN-γ) [3]. The Th1 cytokines, such as interleukin (IL)-12, IL-23, IL-18 and TNF-α, are essential to inhibition of bacterial growth and restrained dissemination [1224]. An increased risk for disseminated NTM disease in humans includes defects in IL-12 [2], IFN-γ [10] or, more recently, interferon regulatory factor-8 (IRF-8) [16]. Some studies have suggested that the expression of immunosuppressive cytokines such as IL-10 and transforming growth factor beta (TGF-β) may be useful to inhibit infection with highly virulent bacterial strains, which are usually susceptible to these cytokines [9]. The active Th1 response and reduced Th2 responses in infected humans or mice protect the body [23]. Although type I IFNs are important to reducing the Th1 immunity during M. tuberculosis infection [25], no studies of type I IFNs during NTM infection have been conducted. Researchers have demonstrated a significant role of CD4+CD25+ regulatory T cells, which are associated with Th1 responses, during M. tuberculosis infection [27]. However, it is necessary to further investigate the function of CD4+ CD25+ regulatory T cells during NTM infection.
Most nosocomial NTM outbreaks are caused by rapidly growing mycobacteria [78], including M. vaccae. M. vaccae was first isolated and named by Bönicke and Juhasz [6] in 1963 using the Latin word for cattle (vaccae) because of its strong connection with cows. M. vaccae has been reported as a cause of clinical diseases, such as bovine nodular thelitis, in cattle [2232]. Moreover, this organism was found to be connected with soft tissue infections and pulmonary disease in four people at the University of Texas MD Anderson Cancer Center (Houston, USA) [15]. Studies have shown an immuno-protective effect of heat-killed M. vaccae on M. tuberculosis-infected mice. Heat-killed bacteria can induce a Th1 response, inhibit the effects of Th2 response, and increase the levels of CD4+T cells and the expression of cytokines [13]. However, few studies have investigated immune mechanisms during a live M. vaccae infection. In this study, we isolated a type of M. vaccae from bovine submaxillary lymph nodes in the Preventive Veterinary Laboratory (Jilin Agricultural University, China) [37]. The goal of this study was to compare the M. vaccae isolated from bovine submaxillary lymph nodes to M. bovis BCG in terms of their infection properties and host immune mechanisms.
Materials and Methods
Bacterial cultures
M. bovis Bacillus Calmette-Guérin (BCG) was obtained from the Preventive Veterinary Laboratory (Jilin Agricultural University). M. vaccae was isolated from bovine submaxillary lymph nodes in the Preventive Veterinary Laboratory (Jilin Agricultural University) [37] and cultured in supplemented Middlebrook 7H9 Broth medium.
Mice
Specific pathogen-free (SPF) female C57BL/6 mice were bought from Beijing HFK Bioscience, China. Mice were used at six weeks of age. All of the experimental procedures complied with the requirements of the Animal Care and Ethics Committees of Jilin Agriculture University.
Mouse infection
The mice were infected with reference strain M. bovis BCG and isolated M. vaccae using intraperitoneal injections of 1 × 106 bacilli per animal. At weeks 2, 4, 6 and 8, the bacterial load was measured by plating serial dilutions of organ homogenates onto supplemented Middlebrook 7H9 agar at 37℃, and the samples were enumerated one week later. A total of five animals were infected for each time point.
Histopathology
The lung samples were fixed in 10% formalin buffer for one week, after which the organs were sliced into 5 µm thick sections and stained using hematoxylin and eosin and acid-fast stains as previously described [25].
Flow cytometric characterization
The cell suspensions were incubated with monoclonal antibodies at 4℃ for 30 min as previously described [2531]. Monoclonal antibodies against CD3 (clone 17A2, rat anti-mouse IgG), CD4 (clone GK1.5, rat anti-mouse IgG), CD8 (clone 53-6.7, rat anti-mouse IgG), CD25 (3C7, rat IgM), CD152 (cytotoxic T-lymphocyte-associated protein [CTLA]-4, cloneUC10-4F10-11, hamster anti-mouse IgG), CD11b (Mac-1, clone M1/70, rat anti-mouse IgG), CD11c (clone N418, anti-mouse IgG), CD45R/B220 (RA3-6B2, rat anti-mouse IgG), and Ly-6C/Gr1 (AL-21, rat anti-mouse IgG) (BD Biosciences). The monoclonal antibody data acquisition and analysis were performed by BD FACSCanto flow cytometry (BD Biosciences), and we examined the samples using a BD C6 flow cytometer (BD Biosciences).
Intracytoplasmic cytokine measurement
The cells were stained as previously described and prepared for intracellular staining [2531]. The cell membranes were permeabilized according to the kit instructions (Fix/Perm kit; BD Pharmingen, USA), after which the cells were stained with antibodies against Foxp3 (MF23, rat anti-mouse IgG), IL-4 (24G2, rat anti-mouse IgG), and IFN-γ (XMG1.2, rat anti-mouse IgG; eBioscience, USA).
Inflammatory cytokine measurement
We collected mouse blood by removing the eyes, placing the blood in centrifuge tubes for 6 h at 37℃, and then centrifuging the samples for 10 min (1,509 × g). The supernatant serum was collected and frozen at -80℃ until analysis for inflammatory cytokines (IFN-γ, TNF-α, monocyte chemoattractant protein [MCP]-1,IL-4, IL-12, IL-10 and TGF-β) using an ELISA kit (BD Biosciences).
Results
Bacterial loads in mice
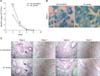
The bacterial load was quantified at weeks 2, 4, 6 and 8 after high-dose (1 × 106 colony-forming unit) intraperitoneal injection. Infection with M. vaccae and M. bovis BCG resulted in severe lung infection at 2 weeks, but the bacterial number gradually decreased and was completely cleared by week 8. The group infected with M. vaccae contained a larger number of bacteria than the M. bovis BCG group (p < 0.001; panel A in Fig. 1).
Histopathology after injection challenge with isolated M. vaccae
The lung histopathology was characterized in mice challenged with M. vaccae and M. bovis BCG (panels B and C in Fig. 1). M. vaccae–infected mice had inflammatory infiltrates with many acid-fast staining bacilli and large aggregates of lymphocytes (panel B in Fig. 1). At week 2, when a larger quantity of bacteria persisted, there was a local inflammatory response in the lungs. By weeks 4 and 6, the moderate-sized lesions had decreased in size. However, at week 8, when M. vaccae and M. bovis BCG strains were cleared, healthy lung tissue was observed (panel C in Fig. 1).
The number of CD3+CD4+ and CD3+CD8+ T cells during M. vaccae infection
The number of CD3+CD4+ and CD3+CD8+ T cells were also determined. There was a strong influx of CD3+CD4+ T cells in M. vaccae-infected mice (panel A in Fig. 2), with the numbers reaching the highest levels at week 4 relative to the other groups (p < 0.01), then declining. However, the M. bovis BCG-infected group did not experience such a rapid decline. There was a rapid influx of CD8 cells, with the highest levels occurring on weeks 6 and 8 of M. vaccae infection compared with the other groups (p < 0.01; panel B in Fig. 2). These results were consistent with the histopathology analysis at week 2, in which we observed many more lymphocytic lesions in M. vaccae- infected mice (panel B in Fig. 1).
The number of CD3+CD4+IFN-γ+ and CD3+CD4+IL4+ cells in the infected mice
To better understand the Th1 response obtained during M. vaccae infection, we examined the number of CD3+ and CD4+ IFN-γ+ cells in the M. vaccae-infected mice. We found that the number of CD3+CD4+IFN-γ+ cells in the M. vaccae-infected mice increased during week 3 and decreased during week 6 (panel C in Fig. 2). Moreover, a strong Th1 response was associated with the emergence of CD3+CD4+IFN-γ+cells initially induced by M. vaccae. In contrast, M. bovis BCG induced a Th1 response that did not decline as quickly.
We also evaluated the Th2 response, which was characterized by the emergence of CD3+CD4+IL4+ cells. As shown in panel D in Fig. 2, a decrease in CD3+CD4+IL4+ cells in the M. vaccae-infected mice occurred during the third week, while the number of cells increased in the sixth week. M. bovis BCG-infected mice did not have an increased Th2 response.
The expression of IFN-γ, TNF-α, MCP-1, IL-4, IL-12, IL-10 and TGF-β in the serum of infected mice
We demonstrated that the serum from the M. vaccae-infected mice contained more IFN-γ starting in the first week of infection and that they had a higher level of IFN-γ in week 4 than the other groups (p < 0.001). Further analysis showed similar results for TNF-α and MCP-1, as well as the production of IL-12. Moreover, IL-4, IL-10 and TGF-β were higher in the M. vaccae-infected mice than in the M. bovis BCG group and the normal group, especially after week 4 (p < 0.001; Fig. 3).
Differences between dendritic cells (DCs) and plasmacytoid DCs (pDCs) in the infected mice
The major producers of type I IFNs are DCs and plasmatoid DC cells [26]. As shown in Fig. 4A, we observed an increased number of DCs during M. vaccae infection to levels higher than were observed in response to M. bovis BCG infection from week 2 to week 4. Similarly, an increased number of pDCs was observed during M. vaccae infection from week 2 to week 8 to levels higher than occurred in response to M. bovis BCG infection (panel B in Fig. 4).
CD4+CD25+Foxp3+ regulatory T cell levels in the infected mice
We evaluated the number of CD4+CD25+Foxp3+ regulatory T cells in the M. vaccae-infected mice. We found evidence of a large number of CD4+CD25+Foxp3+ regulatory T cells in the M. vaccae group (p < 0.001), as well as rapid expression of the cells at week 4 (p < 0.001). A smaller number of these cells was observed in the M. bovis BCG group (p < 0.05; panel A in Fig. 5).
To further investigate the expression of immunosuppressive Treg cells in the M. vaccae-infected mice, we examined mouse CD152 (CTLA-4). The expression level of CD152 can directly or indirectly regulate the level of Treg cells, and it plays a negative regulatory role in the immune response [26]. We found a stronger influx of CD4+CD25+CD152+ T cells during M. vaccae infection from week 1 to week 3 than in M. bovis BCG-infected and normal mice, but these cells declined after 4 weeks of M. vaccae infection in conjunction with a reduced number of CD3+CD4+ T cells (panel B in Fig. 5). In contrast, M. bovis BCG-infected mice contained a large number of CD4+CD25+CD152+ T cells after 4 weeks (p < 0.001). Therefore, the decreased expression of CD152 by M. vaccae-infected mice led to a lack of CD152 signaling.
Discussion
Over the past few decades, researchers have exerted intense efforts to elucidate the host immune mechanisms activated in response to infection with NTM, but have had limited success [1729]. M. vaccae is a fast-growing species of saprophytic bacteria that is widely distributed [6]. It is interesting to note that most of the patients with pulmonary M. vaccae infection had a history of life on cattle farms [15]. However, further studies are needed to investigate the epidemiology, clinical manifestations, and host immune mechanisms of live M. vaccae infection because studies conducted to date have focused on heat-killed M. vaccae. It is known that heat-killed M. vaccae can cause decreased Th2 responses and increased Treg and Th1 responses [1936], which can protect the host against tuberculosis infection. However, to the best of our knowledge, it is currently unknown if these changes occur following infection with living M. vaccae.
In this research, we investigated the host immune responses against live M. vaccae infection. The results showed that the isolated M. vaccae was clearly more virulent in mice than M. bovis BCG, grew much faster in lungs and induced severe lung damage and consolidation, especially during the first 2 weeks of infection. After 2 weeks, the infection and lung pathology slowed due to the protective Th1 response induced by M. vaccae, including a high level of TNF-α and IFN-γ production. Furthermore, M. vaccae inhibited the Th2 response, including lowering the level of IL-4. Previous studies have shown that heat-killed M. vaccae can induce a Th1 response and inhibit the effects of Th2 response [39], which was similar to the results of the present study. However, the Th1 response subsequently declined in conjunction with the emergence of Treg cell response after week 4. There was also decreased production of IFN-γ and TNF-α and evidence of increased Th2 response, which was associated with a high level of IL-4 and differed from previous studies. There are several possible reasons for these differences. Specifically, we utilized live M. vaccae administered intravenously at 1 × 106 bacilli per animal. Additionally, we investigated the effects in healthy mice, whereas previous studies evaluated the immune-protective effects of heat-killed M. vaccae or M. tuberculosis in infected mice that were subjected to multiple immunizations with 107 bacilli via tail intravenous injection [3840]. Therefore, our model reflects those individuals infected with live M. vaccae more clearly.
Earlier studies of M. tuberculosis hypothesized that the type I IFNs were one type of protection for infected animals [18]. A high level of DCs and pDCs, which are type I IFN-rich sources [5], was observed in the M. vaccae-infected group. In the present study, DCs and pDCs showed a significant response to M. vaccae. We also evaluated the T-cell response during infection. CD3+CD4+IFN-γ+ cell numbers increased rapidly, but then declined at the fourth week after infection, whereas we observed a rapid increase in CD3+CD4+IL4+ cells and large production of IL-10 or TGF-β. Some immunosuppressive cytokines such as IL-10 and TGF-β have previously been confirmed to negatively influence the production of TNF and IFN-γ during mycobacterial infection [20]; therefore, we observed low inductions of TNF-α and IFN-γ from the fourth week, which were associated with the emergence of a high level IL-10 or TGF-β production. The function of Treg cells, which are characterized as CD4+CD25+FoxP3+ cells [28], during M. tuberculosis infections is not yet clear because their role has primarily been evaluated during early immune response to such infections [34]. Treg cells that delay T cell priming are present in infected tissue, and mycobacterial immune recovery of patients with HIV infection disease seems to lead to generation of a large number of Treg cells [30]. However, there has been little research on the function of regulatory T cells during M. vaccae infection. The results of the present study indicated that the presence of Treg cells was associated with inhibition of the acquired immune response by a reduction in IFN-γ in M. vaccae–infected mice. Furthermore, we determined the number of CD4+CD25+CD152+ cells in M. vaccae-infected mice, which was associated with the expression of immunosuppressive Treg cells and was down-regulated by week 4. We believe that the decreased expression of CD152 during M. vaccae-infection at week 4 and the coinciding decrease in IFN-γ levels represents a lack of CD152 and fewer activated T cells.
Our study describes the immune processes involved in host defense against infections with M. vaccae. Several different immune responses differed compared to heat-killed M. vaccae infection, and we demonstrated that the M. vaccae isolate was more virulent. Moreover, our study showed several new aspects of the immune response during NTMs infection, which may help improve our understand NTMs and provide a solid basis for treatment strategies.




 XML Download
XML Download