

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
When interacting with a host, viruses may continue to mutate and evolve. RNA retroviruses tend to more easily mutate and evolve than DNA viruses because the RNA polymerases of retroviruses do not have proofreading activity. This could easily lead to the production of more variable viral genomes, thereby forming a large population of variants known as quasispecies [13]. There are various selective pressures aimed at viruses that not only accelerate mutation rates, but also help some rare variants evolve to dominant ones by changing their antigenicity [7]. Increasing attention has been given to viral evolution under immune selective pressures in recent years. For example, the envelope glycoprotein-encoding genes of the human immunodeficiency virus (HIV) have higher mutation frequencies, and their third hypervariable region (the V3 loop) has a mutation frequency as high as 50% [2]. The V3 loop of HIV-1 is closely related to its cell tropism, replication kinetics, and cellular pathogenicity. Mutations in the V3 loop can help viruses escape from attacks from cytotoxic T cells or neutralizing antibodies [1017]. The hemagglutinin glycoproteins of human respiratory Measles virus can also change their antigenicity under immune selective pressures [22].
Avian leukosis virus subgroup J (ALV-J) was the first reported and identified as a new subgroup in England in 1988 [1415]. The gp85 protein, which is encoded by the envelope gene (env) of ALV and located on the surface of virions, binds with receptors on host cells and is the major subgrouping determinant. However, further studies are necessary to define the role of host immune pressures in viral evolution, especially of the gp85 gene. According to a recent epidemiologic study, the gp85 gene of ALV-J mutates much faster than the gp85 genes of subgroups A, B, C, and D. Venugopal et al. [20] compared 12 ALV-J strains isolated in the United Kingdom with the prototype strain HPRS-103 and found that their gp85 amino acid sequence homologies ranged from 92.0 to 98.8%. In the past decade, ALV-J outbreaks have occurred worldwide, causing serious damage to the chicken industry [81923]. In previous studies, we analyzed mutations in eight ALV-J strains that were isolated in China and compared them with strains that were isolated in the United Kingdom and the United States. The results indicated that immune selective pressures may have influenced evolution of the gp85 gene [621].
ALV induces tumors or leukemia in domestic and wild birds. The genetic diversity of ALV has been reported to vary between chicken flocks in different areas and in different years, and even between individuals [681820]. In all of these studies, one random clone of ALV-J gp85 from a bird, or even from a flock, was used as a representative sequence to compare the sequences of strains that were isolated from other flocks or areas in different periods, but the genomic diversity of ALV-J within an individual chicken was not considered. In this study, the diversity of the gp85 gene of ALV-J variants in different organs of the same experimentally infected chicken was analyzed and compared to that of the original inoculum.
Materials and Methods
Virus strain and bird inoculation
A well characterized ALV-J strain, NX0101, was used in the study. This strain was originally isolated from a meat-type parent breeder, white chickens in Ningxia autonomous region, China, in 2001 [6], and its gp85 sequence was registered in GenBank under accession No. DQ115805 (National Center for Biotechnology Information, USA). The stain was cloned via its infectious clone [25], then passaged five times in chicken embryo fibroblast cultures to create a viral stock before use. We inoculated 5-day-old embryos with 0.1 mL (equivalent to 3,000 50% tissue culture infective doses) of the virus through the yolk sac. Approximately 20 chickens were hatched and raised for more than 7 months. All animal experiments were performed according to the guidelines of, and approved by, the Shandong Province Animal Ethics Committee (China).
Isolation and identification of ALV-J in tissues
Tissues were collected from the liver, lungs, and kidneys of the same bird. Each collected tissue sample was fully homogenized on ice in 1 mL phosphate-buffered saline, then immediately centrifuged for 5 min at 4℃ at 10,000 g/min. Each supernatant was subsequently filtered through a 0.22 um filter and inoculated into DF-1 cells to isolate ALV-J variants. The culture supernatants were then replaced with Dulbecco's Modified Eagle Medium (Gibco, USA) containing 2% fetal bovine serum after 2 h, after which they were maintained for 7 day at 37℃. The infected DF-1 cell monolayer in each plate was passaged once or twice, then incubated for another 5 day after each passage. Subsequently, DF1 cells were fixed for 5 min with an acetone:ethanol mixture (3 : 2, v/v). An indirect immunofluorescence antibody assay (IFA) was then conducted using the anti-ALV-J monoclonal antibody JE9 aimed at gp85 protein at 37℃ for 45 min [16], followed by goat anti-mouse secondary antibody for 45 min. The supernatants were saved as virus stocks, and the cells were used for genomic DNA extractions as described below.
Polymerase chain reaction (PCR) amplification of the gp85 gene of ALV-J isolated from different organs
Genomic DNA was extracted from the different tissues and used as the template for PCR. One pair of primers for amplification of the gp85 gene was designed: forward primer, 5′–CTAGCGCCTGCTACGGCGGTGACCC–3′; and reverse primer, 5′–GCATTTCTGACTGGGCACCCTG–3′. The PCR conditions were as follows: 95℃ pre-denaturation for 5 min, followed by 33 cycles of denaturation at 95℃ for 30 sec, annealing at 57℃ for 30 sec, extension at 72℃ for 1 min, and then final extension at 72℃ for 10 min. PCR products were identified by 0.8% agarose gel electrophoresis, after which DNA bands of the correct size were purified using the OMEGA Gel Extraction Kit, cloned into the pMD18-T vector (Takara Bio, Japan), and transformed into Escherichia coli strain DH5α (Takara Bio). More than 10 clones were sequenced for each PCR product. Next, 10 ALV-J gp85 sequences from each organ were chosen for homology comparisons using the DNAStar Lasergene 6.0 software (DNASTAR, USA). The gp85 gene was also amplified and sequenced from DF-1 cells which inoculated with the original NX0101 stock.
Results
Lesions and identification of ALV-J variants in different organs of a dead chicken
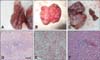
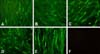
At 28 weeks, a white meat-type parent breeder died of an ALV-J infection. The bird had persistent viremia, and exhibited temporary antibody responses to ALV-J, which resulted in extreme malnutrition. At necropsy, myelocytoma-like tumors were detected in the liver, lungs, and the left kidney. The liver was significantly enlarged and full of small, white tumor nodules that were distributed throughout the entire organ, as occurs in most field cases (panel A in Fig. 1). The lungs and the left kidney also had tumor nodules (panels B and C in Fig. 1), but there were no apparent tumor lesions in the right kidney, which was of the normal color and size (panel C in Fig. 1, right). The left kidney was 5- or 6-fold larger than the right one, and contained several gray-white tumor masses (panel C in Fig. 1, left). Histopathology showed diffuse infiltration of myeloid-like cells in sections of the liver, lungs, and right kidney (panels D-F in Fig. 1). An IFA indicated that DF-1 monolayers inoculated with filtrates from each organ reacted with the ALV-J-specific monoclonal antibody JE9, as indicated by green fluorescence in the cytoplasm and a lack of fluorescence in the cell nucleus (Fig. 2).
Homology comparisons of the gp85 sequences of 10 clones from the same and different organs
Ten clones of PCR products from DF-1 cells that were inoculated with extracts of each organ were sequenced for the gp85 homology comparison. The ALV-J diversity within the same birds is shown in Table 1, which lists the average homologies (ranges) of 10 gp85 clones of each organ and between organs. As indicated in Table 1, the average homologies of the 10 clones from each organ ranged from 98.7 to 99.3%, but the lowest homologies within the same organs were 96.7% (left kidney), 97.4% (right kidney), 97.4% (liver), and 97.6% (lungs). These findings demonstrate the diversity of the ALV-J gp85 gene, even from clones that were isolated from the same organ.
Table 1 also shows that the diversity of isolates from different organs was greater than that from the same organ, and that the lowest homology (94.9%) was observed for ALV-J isolates from the left kidney and lungs. Sequence differences were also compared between the left tumor-bearing kidney and the normal right kidney, which demonstrated that the average homology and the lowest homology between the two kidney isolates were always higher than that between isolates from each kidney and the liver or lungs.
To understand whether the diversity arose during replication in DF-1 cell cultures or in the chicken, 10 clones from DF-1 cells that were infected with the NX0101 stock were also compared. Homology analysis indicated that the homologies ranged from 99.1 to 99.9% among these 10 clones, and that they were 99.2% to 99.9% identical to the original reference strain, NX0101 (GenBank No. DQ115805), even after five passages in DF1 cells. These results suggest that the gp85 gene was relatively conserved during the cell culture passages. However, the gp85 gene started to mutate when ALV-J replicated in the infected chicken. The gp85 gene homology between the original inoculum and the isolates from different organs ranged from 94.1 to 95.8% (liver), while homologies of 94.9% to 96.0% and 93.7% to 95% were observed in the kidneys and lungs, respectively (Table 1).
Comparisons of NS and S mutations in the gp85 gene among clones from different organs of the same bird
As shown in Table 2, the ALV-J isolates from the infected bird had many more mutated sites in the gp85 gene than the original NX0101 inoculum, and their NS/S ratios were significantly higher than those of NX0101. The NS/S ratios that were calculated from 10 gp85 clones of the left kidney, right kidney, liver, and lungs were 6.38, 5.86, 4.41, and 4.00, respectively, while it was only 0.75 for the NX0101 stock (p < 0.05). These results suggested that some selective pressures in the infected chicken influenced the viral mutation rate and, hence, viral evolution during viral replication, when compared with replication of the NX0101 stock in DF-1 cells, thereby promoting diversity of the virus.
The results also showed that ALV-J isolates from both kidneys, whether tumor-bearing or not, had significantly higher NS/S ratios than those of the liver and lungs of the same bird (p < 0.05), although the total number of mutated sites did not differ among them (Table 2). These findings indicate that environment or types of cells in the kidneys placed stronger selective pressures on ALV-J than those of the other organs, such as the liver or lungs, regardless of the presence of tumors.
Analysis of mutational sites in gp85 related to viral evolution in different replication environments
When compared to the 308 amino acid reference sequence of gp85 from ALV-J strain NX0101 registered in GenBank, 27 NS sites were observed in the 50 clones (Table 3). Most of these sites existed in clones that were isolated from the four organs, while only six mutated sites were found in the 10 clones isolated from DF-1 cells that were directly infected with the NX0101 stock. These findings indicate that more mutations occurred in the infected chicken. Nine amino acid residues (61, 68, 88, 134, 168, 189, 205, 212, and 219) in gp85 were mutated in the 40 clones from the four different organs, but no mutated sites were found in the 10 clones from the DF-1 cells that were directly infected with the NX0101 stock, suggesting that these mutated residues may provide the ALV-J variants with some growth advantages that enabled them to out-compete the variants in the original inoculum.
To determine if there were some organ- or tissue-specific selective pressures on viral evolution, the mutational sites in gp85 were compared among organs. When compared with the reference NX0101 sequence, all 20 clones from both kidneys (with or without tumors) had the same amino acid substitutions in six residues (68, 114, 147, 150, 151, and 189); however, not all clones from the liver and lungs demonstrated the same changes. Additionally, there were no mutations in these sites in the 10 clones that were isolated from the DF-1 cells that were infected with the original NX0101 stock (Table 3). Moreover, five of the six sites were located in hr1 (residues 104–156) and hr2 (residues 180–202). The gp85 mutational sites in the left (tumor-containing) and right (tumor-free) kidneys were used to identify any mutational sites that were associated with tumor cells. The only difference between them was that one of the 10 clones from the left kidney had three amino acid mutations (N63D, T101V, and S208R) that were not found in the 10 clones from the right kidney. Interestingly, the same amino acid alterations were detected in some clones of the tumor-bearing liver and lungs.
Discussion
The defining characteristic of RNA viruses, especially retroviruses, is lack of an RNA polymerase proofreading mechanism, which causes instability and increased diversity in their genomes [345]. ALVs, which belong to Retroviridae, are likely to have a high mutation rate and genomic diversity. ALV genomes contain the gag, pol, and env genes and the long terminal repeat (LTR). The gag and pol genes are highly conserved among different subgroups. The LTR is variable, but not related to subgrouping. Distinctions among ALV subgroups primarily depend on the env gene, and especially the gp85 gene [1681824]. Usually, subgroup J viruses exhibit more mutations in the hr1, hr2, and vr3 regions of gp85 than other subgroups, especially in hr1 and hr2, which were hypothesized to be major regions influenced by immune pressures, as demonstrated by their NS/S ratios and antigenic determinants related to virus neutralization [620]. Since ALV-J was first isolated and identified in 1988 [11], many studies conducted worldwide have examined its molecular epidemiology [68911141518192024] to demonstrate the diversity and evolution of the gp85 gene in ALV-J variants isolated from different flocks in different areas at different times. In these reports, only one clone of any ALV-J isolate representing an infected bird, flock, or farm in a given period was sequenced; thus, the genomic diversity of ALV-J strains circulating in the same bird or flock was not considered. However, our laboratory detected such diversity in the gp85 gene of an ALV-J field isolate when more clones were selected for sequence comparisons [12].
In the present study, one chicken that suffered from viremia and exhibited ALV antibodies died suddenly. Necropsy revealed typical myelocytoma tumor lesions in the liver, lungs, and the left kidney, but not the right kidney. Genomic diversity was further demonstrated by sequence comparisons of ALV-J clones that were isolated from different organs of a tumor-bearing bird that was infected as an embryo with the well-known ALV-J strain, NX0101. The results indicated that there was not only a large difference among the re-isolated viruses and the original inoculum stock, but also a wider variation among clones within the same organ or among different organs. The minimum homology of gp85 was only 94.9% between the left kidney and lungs of the same bird. Such a low homology is sufficient for molecular epidemiology studies to separate two ALV-J strains in phylogenic trees. This suggests that any one clone selected from an isolate from any organ of a chicken may not be representative of the viruses that circulate in a flock. Similar gp85 sequence comparisons were also conducted for ALV-J variants that were re-isolated from NX0101-infected birds that lacked tumors. The range of ALV-J gp85 diversity was found to be highly dependent on the infection duration and antibody selective pressure, but less affected by the presence of tumors in this study (data not shown).
In this study, 40 gp85 clones of ALV-J variants from four different organs were sequenced and compared. The right kidney (without tumors) could be used as an ideal control for the other three tumor-bearing organs (the liver, lungs, and left kidney). The gp85 homology of 98.1% (96.6%–99.8%) between the left and right kidneys was higher than that between the left kidney and the liver (96.1%, 95.8%–99.6%) or lungs (95.8%, 95.4%–99.9%), which implies that organ- or tissue-specificity played a more important role than the presence of tumor cells during ALV-J evolution in the infected chicken.
The ratios of NS to S mutations of the gp85 gene were analyzed to determine whether there is any selective pressure that influences viral evolution. It is generally recognized that an NS/S ratio greater than 2.5 indicates that viral genes are under selective pressure, such as immune selective pressure; otherwise, changes in nucleic acids likely result from random mutations [20]. In this study, the NS/S ratios of the gp85 gene from all four organs were higher than 4.0, indicating that there are some selective pressures on viral evolution, such as immune selection pressure or tissue-specific tropism. Moreover, some organ- or tissue-specific positive selective pressures on ALV-J diversity and evolution were observed, as indicated by the fact that the NS/S ratios of the left and right kidneys were closer to each other and significantly higher than those of the liver and lungs. Analysis of the mutational sites in gp85 indicated that each of the 10 clones from the left and right kidneys had very similar amino acid alterations at most mutational sites, but that they differed from those of the liver and lungs, which strongly supports the hypothesis that ALV-J variants that were re-isolated in DF-1 cell cultures of tissue samples from four different organs of the same bird were released from normal histiocytes of each organ, and that organ- or tissue-specificity had a greater influence than tumor cells on ALV-J evolution. Based on the above results and analysis of the genomic diversity of ALV-J isolates from different organs of the same bird, we suggest that more clones from different organs or tissues of infected birds should be sequenced and compared to better understand ALV-J evolution between different flocks and at different times in the field.



 XML Download
XML Download