

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Ticks transmit more pathogens than any other group of blood-feeding arthropods worldwide, affecting humans, livestock and domestic animals [12]. Ixodid ticks of the genera Rhipicephalus, Dermacentor, Ixodes, and Amblyomma are the most important vectors for Rickettsiaceae, including pathogens known to cause disease in humans [2728].
Tick-borne diseases (TBD) caused by Anaplasma (A.) phagocytophilum, Rickettsia (R.) rickettsii, Ehrlichia (E.) chaffeensis, E. canis and other rickettsial pathogens affect humans and/or wild and domestic animals. These pathogens are maintained in natural cycles involving wild mammals and hard-ticks or domestic cycles [11]. The distribution and epidemiology of vector-borne diseases reflects the geographic distribution and seasonal activities of the vectors, reservoirs, animal care, and human behavior. Tick-borne diseases are commonly encountered in medical and veterinary clinical settings, and have increased in recent years, gaining more attention from physicians and veterinarians. Early signs and symptoms of these illnesses are notoriously nonspecific, mimicking benign viral illnesses, which often leads to misdiagnosis and treatment failure with unfortunate, and sometimes severe outcomes [9]. With the application of molecular methods, new species, strains, and genetic variants of microorganisms are being detected in ticks worldwide, and the list of potential tick-borne pathogens continues to grow [30]. The case fatality rate ranges from 4–25% in patients with rickettsioses, 3% in ehrlichioses and 1% in anaplasmoses if treatment is not received [39].
Screening ticks for disease-causing pathogens provides useful epidemiological information on their distribution and the prevalence of pathogens that pose veterinary and medical health risks [6]. This study was conducted to estimate the frequency of Ehrlichia, Anaplasma and R. rickettsii in ticks collected from forests, eco-tourist parks, and hosts, including humans, to determine the geographic distribution and potential exposure to humans and domestic animals in Mexico.
Materials and Methods
Sample collection
Ticks were collected from 1997–2013 from forests, eco-tourist parks, vegetation and wild animals in 22 states of Mexico. In the grass, we used a tick drag consisting of a 1.0 m wide × 1.5 m long white cotton cloth attached to a wooden dowel and a rope attached to each end of the dowel as described by Chong et al. [37]. Drags were pulled at a slow to moderate walking pace behind the collector over vegetation and ground cover. Additionally, ticks were collected from various species of live wild caught rodents and other small mammals by grasping the mouthparts with a fine tweezers and gently pulling them off the host, after which they were placed in 1.5 mL cryovials with 70% ethanol. Ticks were stored until identification and extraction of DNA in the Emerging Infectious Diseases Lab in the National Medical Center XXI Century. Identification was based on their morphological characteristics using relevant taxonomic keys [151737]. A data form that included the collection site, date of collection, species, and stage of development was prepared for each collection.
DNA preparation
DNA from the tissues of ticks was prepared individually. Briefly, ticks were disinfected by immersion in 70% ethanol solution for five minutes, rinsed with sterile water, and then dried on filter paper. Adult ticks were dissected using an aseptic technique and a sterile scalpel to obtain intestinal tissue, while whole larvae and nymphs were used for DNA extraction. Pools were made by stage of development and species after identification. Overall, DNA from 68 individual adult ticks, as well as pools of adults (2 ticks, 286 pools), nymphs (3–4 ticks, 103 pools) and larvae (4–5 ticks, 20 pools) were analyzed. DNA extraction from ticks was performed with a DNeasy Tissue Kit (Qiagen, Germany) according to the manufacturer's instructions. The pathogens detected in the pools were expressed as the percentage and minimum infection rate based on the assumption that each polymerase chain reaction (PCR)-positive pool contained a least one positive tick [13].
PCR for tick identification
We selected 5% of the ticks (51 ticks) from small wild mammals and vegetation (7 adults, 32 nymphs, 12 larvae) for molecular identification using the 12S rRNA gene to verify the accuracy of the morphological identification and then stored species by stage of development. Specifically, larvae (4 pools), nymphs (11 pools), and adults (7) were stored in 1.5 mL tubes containing 70% ethanol until assay.
PCR for R. rickettsii, A. phagocytophilum, E. chaffeensis and E. canis: Tick DNA was amplified with species-specific primers as previously described. The 16S rRNA gene fragment was amplified from all the targeted genes, except R. rickettsii, for which the glta2 and 17kDa genes were amplified. PCR was performed as previously described [3033] using primers Ge3A-Ge10-Ge9F for A. phagocytophilum (16S rRNA gene), HE1-HE3 for E. chaffeensis (16S rRNA gene), ECA-HE1 for E. canis (16S rRNA gene), CS78-CS323 (gltA gene) and Tz15-Tz16 [33] (17 kDa protein-encoding gene) for Rickettsia spp. Positive controls consisted of DNA form the spleens or hearts of Peromyscus leucopus naturally infected with E. chaffeensis and A. phagocytophilum, DNA from a dog infected with E. canis [303233], and DNA from kidney tissue of a patient infected with R. rickettsii. PCR reaction mixtures consisted of 2 pmol of each primer, 200 µM concentration of each deoxynucleoside triphosphate, PCR buffer, 1 U of Taq DNA polymerase (Invitrogen, Brazil) and 50–100 ng of sample DNA for each PCR in a 50 µL mixture reaction.
The PCR products of expected sizes were purified using a QIAquick Gel Extraction Kit (Qiagen, USA), after which forward and reverse nucleotide sequences were determined using a DNA sequencer (Applied Biosystems, USA). Sequence data were collected using the Chromas Lite software (ver. 2.1.1; Technelysium, Australia), and sequences were aligned using MEGA 5.0 [35] and compared with genetic sequences using BLAST (National Center for Biotechnology Information, USA).
Analyses of results were performed using a chi squared or Fisher's exact test, and the odds ratio calculated with 95% confidence intervals. A p value < 0.05 was considered significant. All analyses were conducted using the Epi Info program ver. 6 (Centers for Disease Control and Prevention, USA).
Results
A total of 1,107 ticks were collected 641 ticks from host, while 466 ticks were collected from forests, grass and herbaceous vegetation. The ticks on hosts were collected from nine humans (38 ticks), 148 domestic dogs (262), three deer (8), 183 mice (229), one turtle (6), four sheep (5), two goats (3), 11 cattle (83), two toads (3), three horses (4), and one rabbit (2). The 101 sites include national parks, ecotourism parks, and recreational sites distributed in 22 states of Mexico (Table 1). A total of 1,107 ticks, including 16 species belonging to six genera were collected (Table 2). Rhipicephalus sanguineus was the most frequently collected (43.4%), followed by Amblyomma cajennense (12.2%), Amblyomma maculatum (10.9%), R. microplus (9.0%), Amblyomma dissimile (6.1%), Ixodes (I.) texanus (1.3%), R. annulatus (2.0%), Amblyomma americanum (1.5%), Dermacentor (D.) nitens (1.8%), I. scapularis (1.5%), D. andersoni (1.3%), and Haemaphysalis (H.) leporispalustris (1.3%), while the remaining species only accounted for 1.4% (Fig. 1).
Of the 38 ticks collected from humans, (6.3%) of Amblyomma cajenennse were positive for A. phagocytophilum and (10%) of D. variabilis were positive for R. rickettsii, while no R. sanguineus were positive for the selected tick-borne pathogens. A total of 466 ticks collected from vegetation were combined in 158 pools. Of these, 13 were positive for A. phagocytophilum (3.8%), while six were positive for E. chaffeensis (3.8%), five for E. canis (3.2%) and two for R. rickettsii (1.3%).
A total of 603 ticks were collected from various wild and domestic animals and placed in 281 pools. Assuming one positive tick/pool, E. canis (12.8%) was the most commonly detected tick-borne pathogen by PCR, followed by A. phagocytophilum (11.0%), R. rickettsii (2.9%), E. chaffeensis (2, 0.8%), and R. amblyommii (0.4%) (GenBank accession No. KP844658, KP8446589, KP84663, KP84664 and KP84665).
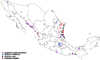
A phylogenetic tree that included the selected tick-borne pathogens was constructed (Fig. 2). A map depicting the distribution of selected tick-borne pathogens detected by PCR from ticks collected by tick drag, humans and wild/domestic animals is shown in Fig. 3. A. phagocytophilum was the most frequent tick infection in the northwest (Table 3). We also identified the following associations: A. phagocytophilum infecting Amblyomma cajennense, Amblyomma dissimile, Amblyomma maculatum and D. variabilis; E. canis infecting Amblyomma cajennense, Amblyomma dissimile, Amblyomma maculatum, I. scapularis and H. leporis-palustris; R. rickettsii infecting D. nitens; and E. chaffeensis infecting Amblyomma cajennense.
Discussion
Since 1947, R. sanguineus has been reported as the tick vector that transmitted R. rickettsi in Mexico [4], while in the United States D. variabilis and D. andersoni tick are the competent vectors for the same pathogen [29]. In 2002, R. rickettsii was identified in R. sanguineus ticks from eastern Arizona as the tick-borne vector of Rocky Mountain spotted fever (RMSF) [10]. In South America, Amblyomma spp. has been reported to transmit other Rickettsia sp. [242627]. In the last ten years, an epidemic outbreak of RMSF has been reported on the northern border of Mexico and into 30 states within Mexico [34],
The tick species reported to transmit R. rickettsii in Mexico include R. sanguineus, D. variabilis and Amblyomma imitator [428], while H. leporis-palustris has been described as a potential vector in northwestern Mexico [37].
We found that R. sanguineus was the most frequently encountered tick, accounting for over 52.2% of all ticks examined in all regions of Mexico. These findings are consistent with the fact that R. sanguineus is considered a global tick capable of transmitting pathogens such as R. rickettsii, Ehrlichia species and even Leishmania infantum [2232731]. The existence of co-infections in individual R. sanguineus adults and nymph ticks was important, and it is known that more than one pathogen may coexist in a single vector [7].
Although the role of R. sanguineus in the transmission cycle of R. rickettsii, the agent of spotted fever rickettsia in humans, has been known in Mexico since the 1940s [34], R. rickettsii was not identified in R. sanguineus in the United States until 2002, when it was identified as an important vector RMSF in eastern Arizona [10]. Our results are similar to reports from Panama, Argentina, and Brazil, where R. sanguineus is a competent vector for most of the tick-borne rickettsial diseases pathogens [824], and was found as the tick most frequently infected with these pathogens.
We found that A. phagocytophilum and E. canis were the most commonly observed TBD pathogens infecting ticks in Mexico, whereas E. chaffeensis and R. rickettsii were less frequent. A previous study in northeastern Mexico reported a prevalence of 50% Rickettsia spp. infection in ticks of the Amblyomma species [24]. Although this frequency is higher than that observed in our study, we performed a more thorough collection, with 16 different tick species. In Texas, Amblyomma cajennense ticks collected from people were found to have multiple infections, including Borrelia spp., Ehrlichia spp., and Rickettsia spp., while Amblyomma maculatum was infected with Ehrlichia spp. and Rickettsia spp. [41].
We report here the presence of A. phagocytophilum, the agent of human granulocytic anaplasmoses, in Mexico. The disease was recognized in humans in the United States in 1994, and subsequently in Europe in 1995 [28]. The Ixodes group is predicted to be present in all of the central and northeastern states of Mexico [13151722]. The main distribution predicted for Amblyomma cajennense is the northeast coast of Mexico, which is characterized by lowlands and warm temperatures. In contrast, Ixodes species are predicted to be present primarily in all of the northern states, which are characterized by high altitudes with a temperate climate and vegetation [1322]. In the United States, I. scapularis and I. pacificus have been found to be the principal vectors for A. phagocytophilum [8].
The detection of the agent of human monocytic ehrlichiosis (E. chaffeensis) in R. sanguineus, Amblyomma americanum and Amblyomma cajennense ticks could indicate that it plays a role in transmission in Mexico. The disease was reported for the first time in Mexico in a patient from Yucatan, who was diagnosed serologically by immunofluorescence assay [14]. In our study, we collected in Yucatan only from R. sanguineus ticks that contained E. canis; however, our results indicate that this infection may occur in other areas in Mexico, including states from the south and north, where the competent vector is predicted to inhabit [13151722].
Amblyomma imitator ticks were found on Silvilagus spp. This tick is a potential vector for R. rickettsii and R. prowasekii, which suggests the participation of other mammals in enzootic cycles in Mexico [28]. We found R. amblyommii in an Amblyomma cajennense tick collected in a northeastern state of Mexico, making this the first report of Amblyomma cajennense as a potential vector for the agent in Mexico. R. amblyommii infection was described in Amblyomma cajennense from the Amazonian region in Brazil [1427] and in Amblyomma neumani in Argentina [26]. Although the pathogenicity of R. amblyommii is still unknown, a study in North Carolina in the United States reported that 11 out of 25 Amblyomma americanum ticks were infected with R. amblyommii, and that three out of six patients had antibodies reactive with R. amblyommii [1].
I. scapularis have been reported in the northern and central states of Mexico since 1962 [21]. D. andersoni ticks were collected from dogs in the State of Sinaloa, in northwest Mexico. These ticks have been described not only in new regions, but also in new biotopes. Various proposed explanations include global warming, the impact of land use, and a role of wild fauna in tick dispersion [513]. Until recently, D. andersoni and D. variabilis in North America, and Amblyomma cajennense in South America were the only species of ticks associated with spotted fever rickettsiae on the American continent [29]. However, other species have recently been reported to be associated with this disease, including Amblyomma triste [38] in Uruguay, Argentina and Brazil, Amblyomma maculatum in the United States and Rhipicephalus sanguineus in Brazil and the United States [2540].
The geographic distribution of TBD usually follows the distribution of tick vectors [1639]. In the present study, we documented the presence of tick-borne pathogens from 67 different locations in Mexico, demonstrating that infected vectors are distributed across the country. We found that A. phagocytophilum was distributed in most of the regions studied, R. rickettsii only in coastal Mexico, E. chaffeensis in the central states and E. canis mostly in northern states of Mexico.
These findings extend previous studies in Mexico, which were limited to the search for a few TBD and tick species, and suggest that most of the tick-borne rickettsial (TBR) pathogens are probably endemic in the country. However, more studies are needed to determine which infections these ticks are able to transmit and cause disease [34].
Overall, the results presented herein provide information about potentially pathogenic TBR organisms, and their tick hosts in Mexico, suggesting the presence of previously unrecognized endemic diseases [18]. It is necessary to determine which associations between ticks and pathogens could result in transmission of the agent, or represent a silent zoonotic cycle of the agent. Knowledge of tick distributions may be useful in predicting the epidemiology of diseases associated with particular tick species [182036], and may also provide an opportunity to examine the ecology of emerging or previously undescribed zoonoses for which different ecological determinants of disease transmission may exist [203334].
The distribution of tick-borne pathogens is changing throughout the world, and some of these agents in Mexico may pose an unrecognized health threat. Our results highlight the need for continued survey studies to monitor the appearance of new enzootic diseases, which may represent unknown threats to public health of the populations exposed.




 XML Download
XML Download