

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Severe fever with thrombocytopenia syndrome (SFTS) is an emerging tick-mediated infectious disease that was recently discovered in the rural areas of China [24]. The clinical characteristics of this disease in humans primarily include thrombocytopenia, leukocytopenia, high fever, gastrointestinal symptoms, and imbalanced liver enzyme production [21]. The pathogen responsible for SFTS is the SFTS virus (SFTSV), a novel Phlebovirus in the family Bunyaviridae [21]. This virus contains three RNA genome segments coated with nucleoprotein (NP) that are encapsulated in an 80- to 120 nm diameter glycoprotein viral envelope [2124]. Since the first report of human SFTS in central China, additional cases have been reported continuously in other parts of China, Korea, and Japan [172526]. In 2013, 36 patients were diagnosed with SFTS, and the fatality rate of this disease was 47.2% in Korea [612].
Among viral pathogen species known to cause human diseases, 76.0% also infect other vertebrate animals [1819]. Moreover, a 41.8% seropositive rate against SFTSV has been reported in domestic animals in China, strongly suggesting that animals are major reservoirs of tick-mediated SFTSV transmission [134]. In addition, seropositive shrews and rodents were recently implicated as potential animal hosts susceptible to SFTSV [9]. A study in Korea showed that SFTSV-positive ticks are widespread throughout most provinces, particularly during summer [13]. Moreover, recent evidence has shown that SFTSV acts as a zoonotic pathogen, and further studies of serological prevalence in farming or wild animal species are necessary.
Reverse transcriptase (RT)-polymerase chain reaction (PCR)-based methods have been conducted to detect the SFTSV antigen in blood samples [716]. Moreover, immunofluorescence assays (IFA) are frequently used to detect SFTSV-specific antibodies in the sera of SFTS patients [23]. These diagnostic methods are associated with several disadvantages, such as the need to handle the live virus, which requires special facilities with high-level biosafety equipment. Furthermore, these methods are laborious when applied to large numbers of samples [21]. In-house indirect enzyme-linked immunosorbent assays (ELISA) and double-antigen sandwich ELISA have been developed to detect immunoglobulins against the NP of SFTSV [5].
Here, we report the development of a competitive ELISA (cELISA) based on the competition of SFTSV-specific antibodies in the test serum with monoclonal antibodies (mAbs) against the NP of SFTSV. The NP recombinant protein is suitable for use as a diagnostic antigen as it is a highly immunogenic protein that has been shown to be expressed from the early stage of virus infection in other Bunyaviridae cases [101123]. We applied this technique to field bovine sera and experimentally generated bovine antisera against SFTSV and investigated the correlation and consistency of this assay with IFA. The cELISA presented in this study can be used to detect antibodies against SFTSV in cattle.
Materials and Methods
Cloning and expression of recombinant SFTSV nucleoprotein
For antigen production, the gene encoding the full-length NP protein of SFTSV strain LN3 (GenBank accession No. HQ141612) was expressed in a bacterial overexpression system. The S fragment of SFTSV was chemically synthesized using the Bioneer gene synthesis service (Bioneer, Korea). The 735 bp gene encoding N-protein was amplified using the following primers: SFTS-NP-1 CTCGGAATTCACATGTCA GAGTGGTCC and SFTS-NP-735 CTTCAAGCTTCAGGTT CCTGTAAGCAG. The PCR amplification was performed using a T3000 thermocycler (Biometra, Germany). The NP gene was cloned into pET-30a(+) (Invitrogen, USA) using the BamHI and XhoI restriction enzymes (New England Biolabs, UK), and subsequently transformed into Escherichia coli BL21 (DE3) (Yeastern Biotech, Taiwan) to express 6xHis-tagged fusion proteins. Following induction with 0.2 mM isopropyl β-D-1-thiogalactopyranoside (AMRESCO, USA) for 20 h at 25℃, the bacterial cell pellet was sonicated in chromatography buffer (20 mM sodium phosphate, 500 mM NaCl, 8 M Urea, and 20 mM imidazole, pH 7.4) and purified using Ni-NTA agarose (Qiagen, Germany) [22]. The affinity-purified protein was urea gradient dialyzed prior to use. The recombinant NP protein was solubilized in 8 M urea buffer, which was then exchanged for 150 mM Tris-HCl (iNtRon Biotechnology, Korea) for further use in subsequent experiments.
Immunization with N protein for rabbit polyclonal antibody and cattle positive control sera production
Animal experiments were conducted according to the protocol of the Institutional Animal Care and Use Committee of the Republic of Korea. Rabbit polyclonal antibody and mouse mAbs were generated after four immunizations with recombinant NP. Polyclonal antibodies were produced in rabbits after four injections of 500 µg of the NP antigen mixed with adjuvant at two-week intervals. The NP-specific mAbs were synthesized as previously described [2]. Briefly, eight mice were injected four times with 100 µg of recombinant NP. Following immunization, the mouse spleen cells were collected and fused with myeloma cells to generate mAbs. Among the mouse hybridoma cell lines, 14 clones were selected according to specific interactions with NP using indirect ELISA.
The cattle were immunized with an injection of formalin-inactivated SFTSV. Briefly, 60 mL of 108 TCID50/mL of the SFTSV was inactivated using 0.025% formalin solution for 72 h. The inactivated virus was concentrated fivefold via filtration using a 10K Ultracel (Millipore, Germany). The loss of viral infectivity was confirmed by culture in Vero cells. Two cattle aged 4 months were used to produce positive control sera against SFTSV. Approximately 2 mL of inactivated SFTSV was emulsified in gel-based adjuvant, and the cattle received intramuscular injections every 2 weeks, with a total of three 3 injections. Serum samples were collected 2 weeks after the last antigen injection. The serum used as a negative control in IFA and cELISA was collected from cattle prior to immunization.
IFA
IFA was performed as previously described, with slight modification [23]. Briefly, Vero E6 cells grown to 80% confluency in 150 cm2 were infected with 2 mL of 1 × 103 TCID50/mL SFTSV. After 6 days, the infected cells (~3,000 cells per well) were spotted onto the microscope slides with reaction wells (Paul Marienfeld, Germany). The cells were immediately fixed with a methanol : acetone (1 : 1) solution for 30 min, after which non-specific signals were blocked with 5% horse serum in a total volume of 20 µL per well. The cells were incubated with mouse mAbs, α-NP specific polyclonal rabbit sera or field bovine serum samples for 1 h at room temperature, followed by detection using fluorescein isothiocyanate (FITC)-labeled secondary antibodies. The FITC-conjugated anti-mouse, rabbit, and bovine antibodies (KPL, USA) were diluted to a concentration of 2.5 µg/mL prior to use. The cells were observed using a Nikon TE-2000U fluorescence microscope (Nikon, Japan).
cELISA
cELISA was performed as previously described, with slight modification [8]. Briefly, a Polysorp ELISA plate (Nunc, USA) was coated with 50, 100 or 200 ng/well of recombinant NP. The plate was then incubated for 16 hours at 4℃ to allow antigen binding. After washing with 0.5% Tween 20 (Sigma-Aldrich, USA) in 1× PBS to remove the unbound antigens, the plate was blocked with 5% skim milk and 1% horse serum in 1× PBS. For the serological test, 50 µL of diluted test sera and the same amount of HRP-conjugated mAb (1 : 500) were added to the antigen coated plate and incubated for 90 min at 37℃. The mAb was conjugated with HRP according to the instructions provided with the EZ-LinkTM Plus Activated Peroxidase kit (Thermo Fisher Scientific, USA). After washing the plate six times, 3,3',5,5'-tetramethylbenzidine peroxidase substrate (KPL) was added to develop the color. The reaction was terminated with 1 M sulfuric acid (Sigma-Aldrich). The optical density (OD) of the sample was calculated as follows: the OD at 450 nm was subtracted from the OD at 630 nm to adjust for the background absorbance. The OD values were converted to percent inhibition (PI) values using the following formula: PI =[1 – (OD test sample/OD negative control)] × 100.
Field bovine sera collection
Bovine field serum samples (n = 416) were collected from breeding cattle in the southern part of Korea (Gyeongsang province) from September 2013 to August 2014.
Application of cELISA to various bovine antisera against other bovine viral diseases
cELISA was conducted using antisera against different viruses: bovine viral diarrhea (BVD) type 1 virus neutralization positive control serum, BVD type 2 virus neutralization positive control serum (National Veterinary Services Laboratories, USA), bovine coronavirus, bovine rotavirus, Bovine leukemia virus and Akabane virus (93FMX) (generated in the Animal and Plant Quarantine Agency, Korea).
Statistical analysis
The positive and negative groups were compared by one-way ANOVA and the Mann-Whitney rank sum test, with p values ≤ 0.001 indicating a significant difference between groups (SYSTAT ver. 4.0; Systat Software, USA). Bee swarm and box plots indicating the distributions of negative or positive groups were generated using an online application [14]. Other graphs and receiver operating characteristic (ROC) [27] curves were generated using the Excel graph function.
Results
Production of polyclonal antibody and mAbs using recombinant NP and the generation of SFTSV-positive sera from cattle
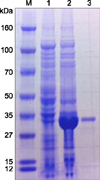
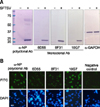
The gene for the recombinant NP was amplified from the commercially synthesized S fragment of SFTSV (GenBank accession No. HQ141612) and cloned into a protein expression vector. A 34 kDa recombinant protein band was well maintained after all purification procedures (Fig. 1). This recombinant NP was used as the antigen to generate polyclonal and mAbs. NP-specific polyclonal antibody was obtained in rabbits after four antigen immunizations, and the titer of the antibody was tested by IFA and Western blotting (Fig. 2). NP-specific mAbs were produced from the spleen hybridoma cells of immunized mice, and the three clones (6D55, 8F31, and 10G7) showed strong interactions in the IFA and Western blot tests; therefore, these clones were used as competitive antibodies to develop the cELISA. A clear band corresponding to the size of NP, which appeared only after SFTSV infection, was observed upon Western blot analysis with all generated antibodies (panel A in Fig. 2). Moreover, IFA using these antibodies showed intense cytoplasmic staining, specifically in SFTSV-infected Vero cells (panel B in Fig. 2).
SFTSV-positive cattle sera were generated from two cattle that were injected three times with inactivated SFTSV. Serially diluted positive or negative serum samples were tested to determine the optimized dilution rate to distinguish SFTSV-positive serum samples from negative samples in IFA. The maximum dilution rate of serum for the IFA at which a non-specific background signal was not observed in the negative control was 1/80 (Fig. 3).
cELISA development and optimization
ELISA plates coated with different concentrations of antigen were used to determine the optimal coating antigen concentration (Supplementary Fig. 1). Among the mAb clones screened based on IgG responses in the IFA and Western blot tests, 10G7 was selected as the competitive antibody for cELISA because this mAb showed stable reactivity after the conjugation of horseradish peroxidase (HRP) protein. The 1 : 500 diluted mAb-peroxidase conjugate yielded the maximum percent inhibition rate in the cELISA (Supplementary Fig. 1).
The immunized cattle serum samples were tested for the cELISA. Both 1 : 2 and 1 : 5 dilutions of the positive serum samples showed higher PI values than the negative serum. However, a 1 : 5 dilution of the positive control sera yielded a large difference in the optical density (OD) between the positive and negative serum samples (Fig. 4). The repeatability of this finding was confirmed by three independent experiments.
The specificity of the SFTSV cELISA was examined using antisera against different types of viruses from cattle. The maximum PI value of laboratory-immunized bovine serum in the cELISA was 93%, while other antisera against different viruses, such as BVD type 1 and BVD type 2, bovine coronavirus, bovine rotavirus, Bovine leukemia virus and Akabane virus (93FMX), yielded PI values of – 18 to 11% (Supplementary Fig. 2). These findings demonstrate that the cELISA is highly specific for the detection of antibodies against SFTSV.
Determination of the cut-off points for IFA and cELISA
A total of 416 bovine serum samples and two positive control sera were tested with the IFA to distinguish negative serum samples from SFTSV. The results showed that 97.4% (407/418) of the bovine serum samples were negative, while 2.6% (11/418) of the bovine serum samples were positive according to the IFA. cELISA was performed using the SFTS-negative group samples, and the mean inhibition percent was 11.3% with a standard deviation of 12.7%. Therefore, we determined the cut-off value to be 49.5% (mean ± 3SD) (Fig. 5). The cELISA and IFA were compared using this cut-off value, and 98.1% (410/418) consistency was observed between results (Table 1). The accuracy of cELISA was demonstrated with a plot of the distribution of PI values obtained from the IFA-negative or positive cattle group (Fig. 6).
Because official standard methods for the antibody detection of SFTSV have not been determined, the cut-off criteria determination for IFA requires further optimization. Therefore, ROC curve was employed to determine the sensitivity and specificity and define the optimal cut-off value for IFA. The ROC curves of four different IFA cut-off values were compared, and the results showed that a 1/80 dilution of the sample strongly supports the result of cELISA. For this dilution, the area under the curve (AUC) was 0.91. While a 1/128 dilution rate as an IFA cut-off point maximizes the specificity and sensitivity compared with other dilution rates, this condition is too strict to detect positive samples above the strict threshold in cELISA. The other IFA cut-off conditions (1/16 and 1/64) were also excluded because they showed moderate AUC values (Fig. 7).
Using a 1/80 dilution as cut-off for the IFA and a PI of 49.5% as a cut-off for cELISA, 0.95% (4/418) of the bovine serum samples were identified as positive, while 97.1% (406/418) of the serum samples were negative.
Discussion
Tick-mediated SFTSV is an emerging infectious virus identified in 2009 that shows 12% lethality in humans [23]. However, the range of host species or virus circulation routes in the environment is difficult to determine because a standard diagnosis method is not available. Therefore, a simple and effective method of detecting SFTSV antibody is needed to identify potential hosts and develop anti-viral strategies. In an effort to detect SFTSV-specific antibodies, in-house ELISA and double-antigen sandwich ELISA (DAS-ELISA) assays were developed [524]. Specifically, DAS-ELISA was used to determine the seroprevalence of humans and animals in China. A multiplexed Luminex-based immunoassay method was also established for high-throughput detection [20]. These newly developed techniques are highly specific and sensitive, but are not commercially available, and some of these methods require special equipment for diagnosis.
Here, we report the development of cELISA using mAb to detect antibodies against SFTSV. The cELISA format was selected as a serodiagnostic test that has value in handling field serum samples because it requires only small volumes of serum for diagnosis and shows species flexibility. Immunostaining and Western blotting were used to detect mAbs with high affinity to SFTSV-infected Vero cells. The results indicated that the generated mAbs were appropriate for use in the development of a cELISA.
A previous study showed that the involvement of domestic animal husbandry is a significant factor in SFTSV infection when compared with outdoor activities or tick exposure in China [15]. Therefore, we collected 416 field bovine sera from farms in Gyeongsang Province in Korea, where 30.6% (11/36) of human SFTSV infections were reported in 2013 [12]. First, we tested the field bovine serum samples using IFA. The bovine serum samples that yielded negative results in the IFA were used as a negative group to determine the cut-off value for the cELISA. All field serum samples and two experimentally immunized bovine sera were examined for an immune response to SFTSV using both IFA and cELISA.
The results showed that 97.4% of the total serum samples yielded negative results in the IFA; therefore, this group was regarded as negative to SFTSV. The cELISA cutoff value was subsequently calculated using the mean inhibition rate of the IFA-negative population ± 3S.D. (cutoff PI = 49.5%). Using this cut-off, 99.8% of the IFA-negative bovine serum samples were also considered to be negative in the cELISA. Although the newly developed cELISA is highly specific, more field animals infected with SFTSV would need to be measured to improve the sensitivity.
SFTSV was first isolated in goats, after which antibodies against SFTSV were detected in the sera obtained from goats at one goat farm in the southern part of Korea (Gyeongsang province) in September 2014 (personal communication with Dr. Lee, Yoon-Hee and Mr. Choi, Jeong-Soo, Foreign Animal Disease Division, Animal and Plant Quarantine Agency, Korea). Serological diagnostic methods are necessary to test large numbers of animals. While the cELISA developed in the present study requires additional tests to validate its performance, the results observed to date are promising. Further studies to adapt this diagnostic technique to a broad spectrum of animal species are currently in progress.





 XML Download
XML Download