

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Trace minerals such as Zn, Cu and Mn are necessary for health in dairy cows because they play important roles in protein synthesis, body metabolism, formation of connective tissue and immune system function [16]. Moreover, they are present in many metalloenzymes involved in the antioxidant process [23]. Oxidative stress is an important pathogenic factor in many diseases [3] that has also recently been found to be involved in the development of lameness in cows [4].
Lameness is a crucial welfare issue in modern dairy husbandry that could result in serious economic losses to dairy producers [27] because of decreased milk yield [5], reduced fertility [721], and increased treatment costs and culling rates [8]. Indeed, lameness has been identified as the third most important health problem in dairy farming following mastitis and infertility [17]. However, it has been shown that supplemental trace minerals (such as Cu, Zn and Mn) may help reduce the incidence of lameness [34].
Regarding trace mineral sources, sulfate salts are readily available and widely used as sources of inorganic trace elements [38], but their efficiency of absorption is very low [26]. Organic trace minerals are reportedly absorbed, stored, metabolized, and transferred more efficiently than in their inorganic forms [29]. Organic forms of Zn, Cu, and Mn have been developed to increase intestinal absorption and mineral bioavailability [40]. Furthermore, Nocek et al. [25] found that supplementation with organic trace minerals can reduce the incidence of lameness. Therefore, this experiment was designed to determine the effects of dietary Zn/Cu/Mn applied as sulfate salts or metal methionine hydroxyl analogs on production performance, different biochemical indicators and indices related to hoof health.
Materials and Methods
Animals, diets and experimental design
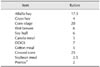
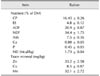
All experiments were reviewed and approved by the Animal Care and Use Committee of Shandong Agricultural University (approval no. 2013-03). Cows selected in this experiment were of similar parity, lactation, and milk production. Before the start of the experiment, the gait score of each cow was determined using a 5-point gait score system (Table 1) [31]. Cows were assigned into two groups of 24 cows each for health or lameness based on their gait score (1 and 2 indicate health and 3, 4 and 5 lameness). Cows in each group were assigned to the following two treatments: (1) control (CON): 50 mg Zn, 12 mg Cu, 20 mg Mn/kg DM as sulfate salts; (2) chelated trace mineral (CTM): 50 mg Zn, 12 mg Cu, 20 mg Mn/kg DM as metal methionine hydroxyl analog (Novus international, USA). The trial lasted for 180 days. Cows were injected with 2 mL of foot-and-mouth disease (FMD) trivalent inactivated vaccine (catalog no. 20130826; Jinyu Baoling Bio-pharmaceutical, China) at day 90 of the experiment period. Cows were housed in individual tie stalls and milked three times a day (3:00 AM, 10:00 AM, and 5:00 PM) while receiving the diets. All cows in this experiment had free access to water and received the same basal diet (Table 2). Basal diet samples were analyzed monthly, and all samples were well above the Nutrient Research Council requirements (Table 3) [26]. Dry matter intake (DMI) was also recorded monthly.
Sampling, measurement, and analysis
Milk yield of the experimental cows was recorded every 10 days, and milk samples were collected for analysis of fat, protein, lactose, and solid non-fat (SNF). Additionally, 15 mL of blood was sampled from the coccygeal vein on day 0, 90, and 180. Serum samples were obtained by centrifuging the blood samples at 1,500 × g for 10 min, then stored at -20℃ for later analysis of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), catalase (CAT) activities, reduced glutathione (GSH), oxidized glutathione (GSSG), malondialdehyde (MDA), metallothionein (MT), procollagen-II N-terminal peptide (PIIANP), C-terminal telopeptide of type II collagen (CTX-II), cartilage oligomeric matrix protein (COMP), interleukin 1 (IL-1), immunoglobulin A (IGA), and Zn/Cu/Mn levels. Serum GSH-Px, SOD, and CAT activities and GSH, GSSG, and MDA levels were measured by spectrophotometry with commercial kits (Jiancheng, China). Serum MT, PIIANP, CTX-II, COMP, IL-1 and IGA were determined using the double-antibody sandwich enzyme-linked immunosorbent assay (ELISA) method with commercial kits (Lengton, China).
In addition, blood samples were collected at day 90, 120, 150, and 180 of the experimental period for analysis of FMD antibody titers. FMD antibody titers (A, O, and Asia I) were measured by the liquid-phase blocking ELISA method with commercial kits (Liquid-phase Blocking ELISA Kit for Detecting Foot-and-Mouth Disease Virus Type A/O/Asia I Antibodies, Lanzhou Veterinary Research Institute, China). FMD antibody titers were log2 transformed before analysis to obtain homogeneity of the residual variance.
At day 0, 90 and 180 of the experiment period, hoof hardness of the apex sole in the heel was tested using a Shore Scale D durometer to measure the ball indentation hardness. Five measurements were made for each cow, and the average value was recorded as the hoof hardness [10]. Hair samples were also collected and analyzed for trace minerals.
For trace mineral analysis, serum samples were thawed before testing. For each sample, 2 mL was digested with 20 mL of concentrated nitric acid and 2 mL of perchloric acid in an Erlenmeyer flask. Samples were digested by heating in an adjustable furnace until the sample became colorless. One gram hair samples were accurately weighed, after which each sample was carbonized in a crucible. Carbonized samples were incinerated in the muffle furnace until completely incinerated, and then dissolved in nitric acid. Digested samples were of constant volume. The levels of Zn, Cu and Mn were analyzed using a flame atomic spectrophotometer (Purkinje General, China) after the volumes of the digested samples were constant.
Statistical analysis
All statistical analyses were performed with SAS (ver. 8.0; SAS Institute, USA) using the cow as the experimental unit. All results were evaluated using a MIXED model including day, lameness, CTM, day × lameness, day × CTM, lameness × CTM, and lameness × CTM × day. Day was included as a repeated measure using an autoregressive covariance structure, and data collected the day before the start of the trial were included as covariates. Significance was declared at p ≤ 0.05, and trends were reported at 0.05 < p ≤ 0.10.
Results
The milk performance data is shown in Table 4. There was no significant difference in DMI, milk yield or compositions between healthy and lame cows. Cows receiving CTM had a significantly lower milk fat percent (p = 0.031). Moreover, cows fed with CTM had numerically higher milk yield and protein yield than those in the CON group (p = 0.102 and 0.103, respectively). However, there were no differences in DMI, protein, lactose, SNF and fat yield between the CON and CTM group.
As shown in Table 5, there was no significant difference between healthy and lame cows for GSH, GSSG, GSH/GSSG, CAT, GSH-Px, PIIANP, CTX-II and COMP. However, lame cows had significantly lower SOD (p = 0.039) and higher MDA (p = 0.031) levels than healthy cows. When compared with healthy cows, lame cows tended to show lower MT (p = 0.087). Cows fed CTM had significantly higher GSH (p = 0.008) and lower GSSG (p = 0.010) values, resulting in a higher GSH/GSSG (p = 0.009) than for those fed the CON. In addition, GSH-Px, SOD and MT were significantly higher (p = 0.011, 0.009 and 0.034, respectively) and MDA, PIIANP and CTX-II were significantly lower (p = 0.007, 0.008, and 0.039, respectively) due to CTM supplementation. CTM tended to decrease serum COMP (p = 0.087). There was no difference in CAT observed between the CON and CTM groups (p > 0.1). Additionally, no interaction between lameness status and CTM was observed in this study for the blood variables tested above.
As shown in Table 6, there was no significant difference in IL-1, IGA, and FMD antibody titers (Type A, O, and Asia I, respectively) between healthy and lame cows (p > 0.1). Additionally, there was no significant difference in the IL-1 and FMD antibody titer- Asian-1 type between CON and CTM groups. However, IGA and FMD antibody titer-O type were significantly higher because of CTM supplementation (p = 0.008 and 0.012, respectively). FMD antibody titer-A type tended to increase for cows supplemented with CTM (p = 0.080).
The levels of Zn/Cu/Mn in serum and hair are shown in Table 7. There was no difference in serum Cu between healthy and lame cows (p > 0.1). However, lame cows had significantly lower serum Zn and Mn than healthy cows (p = 0.007 and 0.08, respectively). Serum Zn, Cu and Mn were significantly higher because of CTM supplementation (p = 0.021, 0.019, and 0.045, respectively). Similarly, there was no difference in Zn and Mn in hair between healthy or lame cows. Lame cows tended to show lower Cu in hair than healthy cows (p = 0.078). Hair Zn, Cu and Mn were significantly higher because of CTM supplementation (p = 0.009, 0.010 and 0.021, respectively).
Lame cows also had lower hoof hardness than healthy cows (Table 8). CTM tended to increase the hoof hardness on day 90 after supplementation (p = 0.085), while significant improvement of hoof hardness was observed on day 180 due to CTM supplementation (p = 0.001), regardless of healthy or lame cows. Finally, CTM affected the gait score of lame cows at the end of the experiment (day 180) (Table 9).
Discussion
No obvious differences were observed in DMI, milk yield and components except milk fat percent between the CON and CTM groups. Similar to one previous study [24], no significant effect was found for organic trace minerals on milk yield and components. Previous studies have shown variable results regarding production response to organic trace mineral sources. A meta-analysis evaluating the effectiveness of supplementation with organic trace minerals found that organic trace minerals increased milk yield relative to control cows [30]. Ballantine et al. [6] and Siciliano et al. [34] also found that supplementation of organic trace minerals to the diet increased milk yield, but had no effect on milk compositions. In this study, cows fed CTM showed significantly decreased milk fat percentage, but the milk fat yield did not differ significantly. These findings suggest that decreased milk fat may be a dilution effect because cows fed with CTM tended to have numerically higher milk yield.
Zn, Cu and Mn play a key role in antioxidant process [23]. Zn plays an important role in the induction and activation of GSH-Px in liver cells, thereby reducing free radicals. In the antioxidant system, Zn, Cu and Mn are components of Zn/Cu/Mn-SOD, which can potentially clear superoxide anion radicals [2037]. Zn can also prevent lipid peroxidation by reducing MDA levels. Moreover, a large amount of MT is produced when levels of available Zn become critical. MT is a potent free radical scavenger, and its major function is to limit oxidative damage [20]. In this study, lame cows had higher oxidant status, but CTM was found to markedly restore the enzymatic (GSH-PX and SOD) and non-enzymatic (GSH and MT) antioxidants levels. Similarly, Osorio et al. [28] found that lame cows had higher levels of oxidative stress than healthy cows. Moreover, supplementation with organic Zn, Cu and Mn was shown to reduce oxidative stress [922]. In contrast, Cope et al. [13] found that there was no effect of feeding organic Zn on serum SOD activity, which may be because only one type of organic trace mineral was provided.
Arthritis is characterized by a limited intrinsic repair capacity and increased cartilage degradation process [18]. Cartilage is composed of cartilage cells and extracellular matrix. Type II collagen is the most abundant protein in cartilage matrix [32]. CTX-II is a degradation product of type II collagen that can reflect the degree of cartilage degradation. Serum COMP, which is a main component of non collagen and the main extracellular matrix protein in cartilage, has been shown to increase in response to arthritis [32]. Interestingly, Aigner et al. [1] found that collagen IIA mRNA is highly expressed in chondrocytes of patients with arthritis, while normal adult cartilage cells did not express it. These findings suggest that the degradation product of collagen IIA levels, i.e., PIIANP, were higher in arthritis patents. However, there were no significant differences in these three arthritis biomarkers observed between healthy and lame cows in the present study. This may have been because the selected lame cows or both healthy and lame cows had very mild arthritis. However, cows fed CTM had lower levels of arthritis biomarkers than those fed with CON, indicating that CTM could reduce the incidence of arthritis.
IL-1 is a cytokine that is produced by macrophages during tissue injury, infection or antigen stimulation. This cytokine is involved in regulation of the immune response and inflammation, and it can increase the host's defensive mechanisms [2]. In this study, no significant difference was observed in changes of IL-1 level between treatments, suggesting that this cytokine was not affected by CTM supplementation. Moreover, another immune marker, IGA, plays a key role in a variety of protective functions via specific receptors and immune mediators [39]. Indeed, IGA can reflect the body's immune function [35]. As shown in Table 6, the value of IGA in cows fed CTM was significantly higher than that in the CON group, indicating that organic trace minerals may strengthen immune function in cows. In addition, cell-mediated immunity has been found to be involved in the clearance of FMD virus from infected animals [11]. Moreover, specific T-cell responses are associated with the induction of anti-FMD virus antibodies [12]. Furthermore, it is known that trace minerals (particularly Zn, Cu, Mn) could activate T-cells and affect antibody responses in the body [15]. In this study, administration with CTM significantly increases the FMD antibody titers in dairy cows. Similarly, organic trace minerals have been shown to increase rabies antibody titer in cows [24]. Shinde et al. [33] also reported that organic Zn could improve the immune response of pigs. Based on these results, there is further evidence that CTM may enhance immune response in dairy cows. Elevated levels of FMD antibody titers varied from different serotypes, which may have been due to several reasons, including the epidemic serotype of the region, secretion mechanism of the antibody, or vaccine quality.
Lame cows have been reported to have lower blood Zn levels than healthy cows [7], which is consistent with the results of the present study. As shown in Table 7, administration of organic trace minerals resulted in increased Zn, Cu and Mn in serum, which may have been due to the better availability of these materials from organic forms. Similar results were revealed in previous studies [19]. In addition, hair trace minerals levels were shown to be the result of long-term accumulation [14]. As shown in Table 7, lame cows had lower Cu in hair than healthy cows, which may have been because of the long-term lack of Cu in lame cows. Administration of organic trace minerals led to increased levels of Zn, Cu and Mn in hair, which further demonstrated that organic trace minerals have better bioavailability.
Keratin, which is the main component of hooves, can improve hoof integrity by speeding wound healing and increasing hardness of the hoof [36]. Zn and Cu are essential minerals of keratin synthesis. As shown in Table 7 and 8, lame cows have decreased hoof hardness, but hoof hardness was significantly improved when they were fed CTM. Moreover, the gait score of lame cows was affected by CTM supplementation (Table 9). Trace mineral deficiencies restricted the keratin synthesis, resulting in lower hoof hardness. In this study, lower levels of trace minerals in the body might have been the main reason for reduced hoof hardness. Furthermore, Cope et al. [13] attempted supplementation with 36 mg organic Zn/kg of dry matter for 14 weeks to improve hoof hardness of cows, but no significant effect was obtained. When compared with our experimental design, we think that organic trace minerals should be fed for a longer time and that a complex of trace minerals (Zn/Cu/Mn) should be used for dairy cows.







 XML Download
XML Download