

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Adipose tissue plays a role in energy storage and thermoregulation, and serves as a source of various hormones including adipokines and cytokines [111236]. Adipose tissue is essential to the absorption of fat-soluble vitamins [35] and for the cell membrane composition [30]. However, constant high-fat diet intake causes obesity by excessively increasing adipogenesis in the body. The increased fat accumulation produces serious negative complications such as increased insulin resistance, arteriosclerosis, cardiovascular diseases, hyperlipidemia and diabetes mellitus [1834,41].
In western countries, many people consume high fat or high caloric diets without regular exercise. Consequently, many pharmaceutical companies have begun investigating drugs targeting obesity. However, several promising anti-obesity drugs have been withheld because they showed unexpected side effects in humans [431].
Artemisia annua (AA), a well-known anti-malaria agent [2329], was recently shown to reduce adipocyte differentiation in many in vitro studies, downregulating the level of peroxisome proliferator-activated receptor (PPAR)-γ, C/EBP-α, C/EBP-γ [22]. However, its effects on adipogenesis have not yet been investigated in animal models.
In the present study, we performed daily oral administrations of AA water extract in a diet-induced obesity (DIO) animal model. AA extract was applied to 3T3-L1 cells, after which the relative protein expressions were compared among various concentrations and times from application. In addition, oil red O staining and western blot were performed in vitro, after which the physiological data and effects on adipogenesis were observed based on the histology and mRNA expression of many related genes to evaluate the anti-obesity effects of the herb AA in an in vivo system.
Materials and Methods
AA extraction
A total of 40 g AA were boiled with 1.8 L distilled water (DW) under 1.5 bar at 80℃. After boiling for 30 min, the extract was fully cooled. The extract was filtered first by paper (185 mm; Advantec, Japan), then by a Nalgene Rapid-Flow Battle Top Filter (0.2 µm-pore membrane; Thermo Scientific, USA). Final AA extract was stored at 4℃.
Animals
Twenty-four adult C57BL/6J mice (mean = 23 g, between 21 and 25 g, 7-weeks-old) were housed at room temperature (22℃) and 60% humidity under a 12-h light: dark cycle (light cycle : dark cycle from 07:00 to 19:00). The mice were divided into four groups, two that were provided a normal chow diet (2018S; Harlan, USA) and two that were given a high fat diet (TD.06414; Harlan). Free access was allowed to water. Every day, 10 mL/1 kg/day AA extract was carefully administered with an oral sonde (0.9 × 50 mm) to half of each food group, while the same amount of DW was administered to the other half. Weight, food and water intake were recorded daily, and blood sugar level was tested once every week under non-fasting conditions. The peripheral blood was collected by cutting the tip of the mouse tail vein. Glucose levels from the peripheral blood were measured using a One Touch Ultra (LifeScan, USA) blood glucose meter with One Touch Ultra test strips (LifeScan). The experiments were performed over 4 weeks and were approved by the Institutional Animal Care and Use Committee (IACUC approval no. SCH15-0001) at Soonchunhyang University.
Tissue processing
Epididymal adipose tissues were removed before perfusion and immersed in 4% paraformaldehyde (PFA). The animals were perfused with 0.1 M phosphate-buffered saline (PBS; pH 7.35) followed by 4% PFA in 0.1 M phosphate-buffer (PB; pH 7.35).
Cell culture
3T3-L1 cells were maintained in high-glucose (25 mM) Dulbecco's Modified Eagle's medium (DMEM; Gibco, USA) supplemented with 10% bovine calf serum (BCS; Hyclone, USA) and antibiotics (penicillin 100 U/mL and streptomycin 100 mg/mL) at 37℃ in a 5% CO2 incubator. For adipocyte differentiation, cells were treated with differentiation inducing media (DIM) containing 1 µM dexamethasone (Sigma, USA), 5 mM 3-isobutyl-1-methylxanthine (Sigma), and 4 mg/mL insulin (Sigma) in DMEM with 10% fetal bovine serum (FBS) at 2 days post-confluence. After 2 days, the cells were cultured in DMEM containing 10% FBS and insulin. Subsequently, the medium was changed every second day.
Oil red O staining
Oil red O staining was performed on adipogenic induction day 8. Briefly, cells were washed twice with PBS, then fixed with 4% paraformaldehyde for 1 h at room temperature. The cells were subsequently washed with 60% isopropanol and dried completely. Finally, cells were stained with oil red O (6 parts 0.5% oil red O powder in isopropanol and 4 parts water) for 10 min and washed with PBS.
Quantitative real-time PCR analysis
Total RNA was isolated from the epididymal adipose tissue using an Ambion PureLink RNA Mini Kit according to the manufacturer's instructions (Ambion, USA). Quantitative real time PCR was performed with SYBR Green dye using an ABI Step One Real Time PCR instrument (Applied Biosystems, UK). For relative quantitation of gene expression, we used the comparative Ct method (2-ΔΔCt). Results were normalized to the control gene (36B4, housekeeping gene, acid ribosomal protein). The sequences of the primers and probes used are listed in Table 1 [1317].
Western blot analysis
Cell extracts were homogenized in lysis buffer (iNtRon Biotechnology, Korea), and protein concentrations were determined with a BCA kit (iNtRon Biotechnology). Lysates were separated with 10% SDS-PAGE and transferred to PVDF membranes (Bio-Rad Laboratories, USA). The membranes were probed with primary antibodies against PPAR-γ, fatty acid binding protein 4 (FabP4), glyceraldehyde 3-phosphate dehydrogenase (Cell Signaling Technologies, USA) and C/EBPβ (Abcam, UK), then incubated overnight. After further washing, membranes were incubated with HRP-conjugated secondary antibody (Vector, USA). The immunoreactive signals were detected based on their enhanced chemiluminescence and recorded in the MicroChemi 4.2 system.
Data analysis
All measurements were performed and analyzed to ensure objectivity. The intensity of the bands generated during western blotting was evaluated based on the optical density measured by transforming mean gray levels using the formula: optical density = log (256/mean gray level) with ImageJ 1.59 software (National Institutes of Health, USA). Lipid droplet size was calibrated per experiment area in the microscope. Dataf are presented as the means ± standard error (SE). Relative mRNA expression levels were automatically measured by real-time qPCR. The differences between means were analyzed using repeated two-way analysis of variance and one-way analysis of variance followed by the Bonferroni post-test and Duncan's new multiple methods to determine differences between experimental groups.
Results
Physiological data
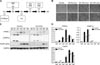
The results showed that all four groups of mice had similar weights at the start of the experiment. Daily weight measurements showed that the high fat diet (HF)/AA group weighed less than the HF/vehicle (Veh) group. This difference was statistically significant starting from day 14, and became more apparent as the experiment progressed. In both normal chow diet (ND)-fed groups, the ND/AA group weighed slightly less than the ND/Veh, but this difference was not significant (panel A and B in Fig. 1). Food intake in both ND fed groups (ND/Veh and ND/AA) did not differ significantly throughout the experiment. For the first two weeks, there were no notable differences in food intake between the HF fed groups (HF/Veh and HF/AA), but food intake in the HF/AA group became relatively lower than in the HF/Veh group after that (panel C and D in Fig. 1). There were no differences in blood sugar levels among groups. Epididymal fat tissues of the mice that received AA (ND and HF) were lower than those of mice that received Veh (ND and HF).
Adipogenic effect of AA water extract in vitro
Adipogenesis was suppressed in the 3T3-L1 cell plate treated with AA water extract, which was the same agent administered orally to the DIO animal model during the experiment. The distinctions between the control plate and the AA treated plate became more obvious at higher concentrations of AA (panel B in Fig. 2). This anti-adipogenic effect was observed by western blotting, which was used to compare the expressions of several factors. PPAR-γ, transcription factor was significantly suppressed on day 3 and 5 in the plate treated with AA. Another transcription factor, C/EBP-β, was also inhibited on days 1 and 2. Moreover, FabP4 which is detected in mature adipocytes, was suppressed on days 3, 5 and 8 (panel C in Fig. 2). The mean gray levels of the all bands were digitized with image J (panel D in Fig. 2).
Epididymal adipose tissue hematoxylin and eosin staining
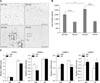
The results of hematoxylin and eosin (H&E) staining demonstrated that the adipocyte sizes in mice administered AA (ND/AA, HF/AA) were smaller than those in mice administered Veh (ND/Veh, HF/Veh). The parts indicated with black squares were magnified. The sizes of the lipid droplets measured using ImageJ and were 1,981 ± 160, 1,177 ± 94, 2,594 ± 258 and 1,232 ± 91 (relative levels) in the ND/Veh, ND/AA, HF/Veh and HF/AA groups, respectively. Both AA-administered ND and HF fed groups demonstrated reduced lipid droplet sizes compared to the Veh-administered ND and HF fed groups (panel B in Fig. 3).
Transcription factor RT-PCR
PPAR-γ, C/EBP-α, C/EBP-β and C/EBP-δ mRNA expressions of ND/Veh, ND/AA, HF/Veh and HF/AA were measured by RT-PCR. There were no differences in PPAR-γ, C/EBP-α and C/EBP-β expressions in the AA-administered groups (ND and HF) or the Veh-administered groups (ND and HF). However, C/EBP-β expression was noticeably more suppressed in HF/AA than in HF/Veh. In the ND-fed groups, this reduction of C/EBP-β expression was found in the Veh-administered group, but the difference was not significant.
Discussion
The number of individuals with obesity has increased gradually in recent decades because of high calorie diets and various social stresses. Many types of anti-obesity drugs and agents have been developed, but these have numerous side effects on the cardiovascular system, gastrointestinal system and central nervous system [4826]. In particular, specific serotonin reuptake inhibitors (SSRIs) and serotonin-norepinephrine reuptake inhibitors (SNRIs) have been investigated and used as appetite suppressants in medical treatments [516]; however, continuous administration of SSRIs and SNRIs causes regaining of weight in obesity patients, despite modifications to these agents to reduce side effects [67]. Therefore, there is demand for natural substances with anti-obesity effects that have low side effects and toxicity [2732].
Many natural substances, particularly AA, have been studied for a variety of applications. AA has been used for extermination of vermin [19] and as an anti-malarial agent [29]. Recent studies have even reported that AA has inhibitory effects on cancer metastasis [38]. Additionally, the anti-adipogenic effects of AA have been investigated and the reduction of adipogenesis by AA in vitro was demonstrated [22]. However, no studies have investigated whether AA has the same effect in vivo. Therefore, we administered a daily dose of oral AA water extract to a DIO animal model in the present study. Our results confirmed the anti-adipogenic effects of AA on both the in vitro and in vivo systems. We measured food intake volume and calories daily and found no differences between the AA-treated groups (ND and HF) and the Veh-treated groups (ND and HF). Although there were no differences in food intake rate, there was a significant difference in body weight between the AA- and the Veh-treated groups. These results indicate that AA water extract may play a primary role in inhibiting adipogenesis rather than suppressing appetite. Interestingly, body weight gain in the HF/AA group was significantly lower than that in the ND/AA group. If this outcome translates to humans, it is expected that AA could reduce body weight gain by decreasing adipogenesis in the abdomens of obesity patients without suppressing their appetite and food consumption.
Western blotting showed that the expression of PPAR-γ, C/EBP-β and FabP4 decreased in the AA treated plate. PPAR-γ is a nuclear hormone receptor that plays a major role in regulating the expression of proteins necessary for development of functional mature adipocytes [915]. C/EBP-β plays a synergistic role in terminal adipocyte differentiation [40], and FabP4 is detected in mature adipocytes [214]. Recent studies have shown that the expressions of these factors also decreased when adipogenesis was suppressed, while they increased when adipogenesis was enhanced [2021]. Therefore, these results indicate that AA inhibits the expression of proteins essential for adipogenesis and/or adipocyte maturation.
H&E staining revealed cell infiltration between adipocytes. The cells may be lymphocytes or macrophages since adipocytes are known to be surrounded by immune related cells in obese animals and humans [3]. It is also well known that measurements of fat accumulation are directly proportional to adiposity [10]. Therefore, overt detection of macrophages indicates excessive adipogenesis. Macrophages, which secrete many types of adipokines and cytokines, are related to the innate inflammation response [24]. Continuous adipose tissue accumulation with macrophages maintains a chronic low inflammatory state, and pro-inflammatory factors such as interleukin-1 and interleukin-6 are continuously secreted [3339]. In particular, interleukin-6 has a harmful effect on the hippocampus, impairing learning, cognition and memory function and leading to dementia [137]. Obesity-related chronic inflammation also contributes to neuropsychiatric symptoms involving depression [2528].
Adipose tissue in HF/AA mice manifested lower cell infiltration than in HF/Veh mice, and the lipid droplet size of HF/AA was less than that of HF/Veh, indicating inhibition of pro-inflammation or inflammation in HF/AA. Additional immunohistochemistry is required to confirm the presence of macrophages; however, the results of the current study indicate that AA exerts an anti-adipogenic effect in the DIO animal model, mitigating adipose tissue accumulation and inflammation. Furthermore, a previous study demonstrated that artemisinic acid isolated from AA significantly attenuated tumor necrosis factor-α-induced secretion of interleukin 6 [22], indicating that AA has an inhibitory effect on the inflammatory state characterizing obesity. Thus, AA is likely to have beneficial effects on the brain of obese individuals, lowering the potential risk of central nervous system disease.
In conclusion, AA water extract decreased body weight gain and suppressed adipocyte differentiation in a DIO animal model, which was reflected in the in vitro results. Additionally, it is expected that AA will improve brain function, particularly on the hippocampus, suppressing the chronic inflammatory state in the DIO animal model. Our study may provide background information for further anti-obesity research.


 XML Download
XML Download