

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Porcine circovirus (PCV; family Circoviridae, genus Circovirus) is a small non-enveloped virus with a diameter of approximately 17 nm and a circular, single-stranded DNA genome [18]. There are two types of the virus: PCV1 and PCV2. PCV2 is often associated with a number of disease symptoms in swine including enteritis, respiratory distress, systemic infection, porcine dermatitis, nephritic syndrome, and reproductive problems. These are collectively known as porcine circovirus-associated disease (PCVAD) [7,14]. In contrast, PCV1 has been shown to be nonpathogenic in pigs under experimental conditions [2,19]. Since characteristics of PCV2 that affect invasion of the immune system enhance the severity of several diseases in swine [7], PCV2-associated diseases are generally recognized as having a significant economic impact on the swine industry worldwide [8]. In addition, PCV2 interacts with other infectious or non-infectious factors (i.e., pathogens, farm type, or swine management strategies) that influence post-weaning mortality in swine [16]. Therefore, detection, surveillance, and preventive measures for PCV2 infection are important for swine producers. The virus is stable [1] and ubiquitous among swine populations [9], which makes eradication difficult. Vaccination thus represents an attractive method for controlling endemic infection and has been shown to be effective for preventing PCV2-associated diseases [5,10]. Several commercial vaccines have been developed and introduced into the swine industry.
PCV2 can be divided into two main subtypes, PCV2a and PCV2b, based on sequence analysis and composition of two open reading frames (ORFs) that encode functional proteins [3,4]. The ORF1 and ORF2 genes produce a replication-associated protein (Rep) for virus replication and an immunogenic capsid protein, respectively [3,13]. It is known that ORF2 has greater nucleotide variation than ORF1 [11]. Therefore, detection of antibodies against PCV2 differs depending on the antigens and detection methods [7,13,14].
Since several vaccines have been introduced into the Korean market, diagnostic laboratories in the country have surveyed for anti-PCV2 antibodies using different enzyme-linked immunosorbent assay (ELISA) kits. Recently, two ELISA kits with different antibody detection systems have been used in Korea. However, no studies have been performed to compare the ability to detect anti-PCV2 antibodies in field pigs resulting from PCV2 vaccines. In the current report, agreement between two commercial serologic assays using sera from field pigs vaccinated with three different PCV2 vaccines is described. Additionally, the prevalence of anti-PCV2 antibodies among pigs in Korea was evaluated.
Materials and Methods
Serum samples from pigs 20, 40, 70, 100, 130, and 160 days of age and sows were collected from farms on which the animals had been vaccinated with the three different PCV2 vaccines (Ingelvac CircoFLEX; Boehringer Ingelheim Vetmedica, USA; Circumvent PCV2 and Porcilis PCV-One; Merck Animal Health, USA). A total of 517 field pig serum samples were studied including 100, 165, and 189 samples from two, three, and three farms with pigs vaccinated with vaccine I, II, and III, respectively, as described in Table 1. The vaccinations were administered according to the recommendations of the manufacturers.
All pig sera were tested using two commercially available ELISA kits: S-ELISA (Synbiotics, France) and M-ELISA (Median Diagnostics, Korea). The ELISA protocols provided by the manufacturers were strictly followed. For the S-ELISA, the controls and serum samples (diluted 1 : 1000) were placed in wells coated with a PCV2 antigen and incubated for 1 h at 37℃. After a wash to eliminate the non-associated components, an anti-PCV2/peroxidase conjugate was added and the plates were incubated for 1 h at 37℃. When there is no specific anti-PCV2 antibody in the serum sample, the anti-PCV2/peroxidase conjugate is free to attach to the antigen in the wells. After a second wash, the enzyme conjugate was visualized by the addition of a substrate that induced a change of color. Following addition of the stop solution, the optical density was measured at 450 nm using an Emax Precision microplate reader (MDS, USA).
For the M-ELISA, the controls and serum samples (diluted 1 : 100) were incubated in wells coated with recombinant PCV2 nucleocapsid protein for 1 h at 37℃. After washing, an anti-pig immunoglobulin G (IgG)/peroxidase conjugate was added and the plates were incubated for 1 h at 37℃. Color development was induced by adding substrate solution to the plate. Optical density was measured at 405 nm using an Emax Precision microplate reader.
The S/N ratio, defined as the optical density of the test serum (S) divided by the optical density of the negative control (N), was calculated for the S-ELISA. Results for the indirect ELISA (M-ELISA) were calculated as the S/P ratio defined as the optical density of the test serum (S) divided by the optical density of the positive control (P). For the S-ELISA, samples with a ratio ≤0.4 were considered positive while ones with a ratio >0.4 were negative for antibodies. For the M-ELISA, samples with a ratio ≥0.4 were considered positive, ones with a ratio between ≥0.3 and <0.4 were false-positive, and ones with a ratio <0.3 were negative. Correlations between data from the two ELISAs were evaluated based on Pearson's correlation coefficient (r) and determination coefficients (R2) calculated with the Statistical Package for the Social Sciences software (ver. 21; IBM SPSS, USA). P values <0.05 were considered significant.
Results
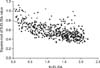
For a total of 517 serum samples, 371 (71.8%) and 472 (91.3%) had positive S-ELISA and M-ELISA results, respectively. The two ELISA assays were fully concordant for 365 positive and 42 negative (including 14 false-positive results obtained with the M-ELISA) samples, indicating 78.7% agreement for the ELISA results. There was a moderate correlation (Pearson r = -0.636, p < 0.001) between the M-ELISA results and square-root transformed S-ELISA results, indicating a linear relationship (y = -0.175x + 0.787, R2 = 0.405; Fig. 1). As shown in Fig. 1, the results for 109 samples were mismatched between the two ELISAs, including four samples with positive S-ELISA results and negative M-ELISA findings along with 105 samples with negative S-ELISA results and positive M-ELISA results. The majority of mismatches were negative for the S-ELISA and positive for the M-ELISA, which were in 0.6 ≥ S/N ratio > 0.4 of S-ELISA and 1.2 > S/P ratio ≥ 0.4 of M-ELISA.
PCV2-specific antibodies were identified in pigs vaccinated with the three different PCV2 vaccines (I, II, and III) using two commercially available ELISA kits. With vaccination, antibody titers against PCV2 were increased in most of the animals. However, the increase and duration of the antibody titer varied depending on the vaccine, presence of maternal antibodies, and vaccination program. Agreement between the ELISAs based on the vaccines was also assessed, and was 76%, 73.9%, or 83% for the pigs given the vaccine I, II, and III, respectively. In addition, results of the ELISAs had a moderate correlation (vaccine I, Pearson's r = -0.602, p < 0.001; vaccine II, r = -0.672, p < 0.001; vaccine III, r = -0.621, p < 0.001) and a linear relationship (vaccine I, y = -0.183x + 0.806, R2 = 0.362; vaccine II, y = -0.191x + 0.815, R2 = 0.451; vaccine III, y = -0.158x + 0.759, R2 = 0.386, p < 0.001) according to the vaccine (panel B in Fig. 2).
Pigs administered I vaccine had different ELISA results at 20 and 70 days of age. The S-ELISA produced negative results at 20 days of age before the antibody levels gradually increased. The M-ELISA produced positive results at 20 days of age before the levels gradually increased after a sudden decline at 70 days of age. For the pigs given vaccine II, the M-ELISA results were positive at all ages and indicated that a seroconversion occurred after the second vaccination. The S-ELISA results showed that antibody levels gradually increased, but the results for some farms (Farm 5) indicated that antibody levels decreased after vaccination. A slight decrease of antibody levels occurred at 40 days of age according to the results of both ELISAs. Pigs treated with the vaccine III showed a gradual increase in antibody levels according to both ELISAs with a significant decrease at 40 days of age based on the S-ELISA findings.
Discussion
Vaccination is an attractive measure for controlling PCV2-associated diseases. Therefore, methods for measuring anti-PCV2 antibodies have been developed and applied to field samples. Serum-virus neutralization, immunoperoxidase monolayer, and indirect immunofluorescent assays have all been widely used to detect anti-PCV2 antibodies [12,17]. These methods can be labor-intensive and time-consuming, and are associated with the risk of virus contamination. In contrast, ELISAs can avoid these problems and perform large-scale diagnostics [17]. The two commercial ELISA kits used in this study involve different antigens and systems. Both ELISA kits involve PCV2 antigen; however, the S-ELISA uses a manufactured version of the antigen while the M-ELISA employs a recombinant nucleocapsid antigen of the Korean isolate. In addition, the M-ELISA is an indirect ELISA while the S-ELISA is a competitive ELISA. Therefore, the results of these two methods are interpreted differently. For example, if there is no specific anti-PCV2 antibody in the serum sample, the second antibody/conjugate of the M-ELISA does not bind to the antigen while the second antibody/conjugate of the S-ELISA is free to attach to the antigen. Using these different ELISA systems, anti-PCV2 antibody concentrations were comparatively evaluated in field pig sera in the present study.
Pearson's coefficient indicated a moderate correlation with 78.7% agreement between the ELISA results. In addition, similar levels of agreement between the ELISAs dependent upon the vaccines was observed with 76%, 73.9%, and 83% for vaccine I, II, and III, respectively. Pearson's coefficients concurred with these findings. It was therefore concluded that the ELISA results based on vaccine type were not significantly different because vaccine I, II, and III use the ORF2 of PCV2 as the antigen.
According to the results of the ELISAs for the field pigs, animals with high levels of anti-PCV2 antibodies at 20 days of age showed a slight increase or maintained antibody levels after vaccination. In addition, slight decreases in antibody levels were observed at 40 days of age in these pigs. This phenomenon was possibly due to maternal-derived antibodies (MDA) that blocked or attenuated responses to the PCV2 vaccination. MDA could interfere with development of an active humoral immune response after PCV2 vaccination [6]. However, this does not mean that vaccine efficacy is reduced because both humoral and cellular immune responses are required for protection [6,15]. In the pigs showing a low level of PCV2 antibody at 20 days of age, there was a significant seroconversion after vaccination.
The M-ELISA results had a broad positive S/P ratio ≥ 0.4 while the S-ELISA had a relatively narrow positive range of this ratio (0 ≤ S/N ratio ≤ 0.4). Therefore, the M-ELISA can be used to detect seroconversion after vaccination more easily than the S-ELISA. Although the results of the two ELISAs revealed similar seroconversion patterns in the field pigs, results for the M-ELISA were more frequently positive than those for the S-ELISA. Data from the ELISAs were controversial at some points depending on different ELISA antigens and practices at each farm. Therefore, further study of samples that produce positive M-ELISA findings and negative S-ELISA results is needed.
In summary, anti-PCV antibodies in pigs were evaluated with two commercially available ELISA kits. Both assays produced comparable results with some exceptions. Based on our findings, the levels of anti-PCV2 antibodies may be used to determine optimal timing of vaccination and verify vaccination compliance of herds.


 XML Download
XML Download