

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Previous experimental evidences have indicated that the mechanisms underlying various types of hepatic injuries are mediated by robust generation of intracellular reactive oxygen species (ROS) [15,33]. Although ROS play normal physiological roles such as second messengers for normal cellular signaling, excessive ROS generation is known to cause cell damage and tissue injury due to an imbalance between pro- and antioxidants [19,38]. Generation of ROS during metabolism and other activities that exceeds the antioxidant capacity of a biological system gives rise to oxidative stress. Many researches have demonstrated that ROS, such as hydrogen peroxide (H2O2), superoxide anions, and singlet oxygen, are involved in many pathological conditions [21,25], and apoptotic cell death could be also promoted depending on the time of exposure and/or concentration of ROS [4,9]. Extracellular H2O2 has been used by investigators to inflict oxidative injury in a variety of cell types including neuronal progenitor cells, HepG2 cells, and hepatocytes at concentration ranging from 100 to 1,000 µM [16,22,29]. Previous studies indicated that extracellular H2O2 increases intracellular ROS levels via multiple mechanisms including loss of intracellular ROS antioxidants such as glutathione (GSH) [23] or decreased mitochondrial membrane permeability followed by mitochondrial ROS release [8]. Therefore, we used H2O2 to induce oxidative injury in primary mouse hepatocytes.
Recent reports suggest that several endogenous and exogenous antioxidants neutralize free radicals and protect the body by maintaining redox balance [27,34]. Antioxidants can perform protective roles against free radicals through a variety of different mechanism including catalytic systems to neutralize or divert ROS, binding or inactivation of metal ions to prevent ROS generation through a Harber-weiss reaction, scavenging and destroying ROS or absorbing energy and electron, and quenching of ROS [28]. Phenolic compounds commonly found in both edible and inedible plants have been reported to have multiple biological or medicinal properties as well as beneficial effects on health [2,17]. Accumulating evidence has indicated that phenolic compounds exert antioxidant effects via a number of mechanisms such as ROS-scavenging activity, regulation of ROS-removing enzymes, antioxidant-synthesizing enzymes, or ROS-forming enzymes [31]. Butylated hydroxyanisole (BHA; 3-tert-butyl 4-hydroxyanisole) is one of the most widely used synthetic phenolic compounds and is regarded as an effective food additive [14]. BHA is known to have various beneficial activities such as antioxidant activity, anti-inflammatory effects, and anticancer potential although some reports suggested that this reagent induces cytotoxicity [26,30,35,39]. However, direct evidence of the effect of BHA on ROS-induced hepatocyte apoptosis has not been obtained. Therefore, we investigated the ability of BHA to protect primary cultured mouse hepatocytes against H2O2-induced apoptosis.
Materials and Methods
Materials
Eight-week-old male ICR mice were purchased from Daehan Bio Link (Korea). All animal management procedures were conducted in accordance with the standard operation protocols established by Kyungpook National University (permission no. KNU 2012-119). A Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Laboratories (Japan). BHA, H2O2, N-acetyl-Lcysteine (NAC), collagen, recombinant human insulin, dexamethasone, Williams' E medium and type IV collagenase were obtained from Sigma-Aldrich (USA). 5-(and 6)-chloromethyl-2',7'-dichlorodihydrofluorescein diacetate, acetyl ester (CM-H2DCFDA) were purchased from Life Technologies (USA). Fetal bovine serum (FBS) was acquired from Thermo Scientific (USA). Antibodies against Bax and Bcl-2 were obtained from New England BioLabs (USA). Anti-PARP-1, goat anti-rabbit IgG, and goat anti-mouse IgG antibodies were supplied by Santa Cruz Biotechnology (USA).
Isolation of mouse hepatocytes
Primary hepatocytes were isolated from mouse liver using the two-step EDTA and collagenase perfusion method. After the mouse was anesthetized, the liver was perfused with Krebs-Henseleit buffer without Ca2 + or SO4 2 - (115 mM NaCl, 25 mM NaHCO3, 5.9 mM KCl, 1.18 mM MgCl2, 1.23 mM NaH2PO4, 6 mM glucose, and 0.1 mM EDTA) through the hepatic portal vein to remove the blood (flow rate of 7~9 mL/min for 5 min). The liver was then perfused with Krebs-Henseleit buffer without Ca2 + or SO4 2 - containing 0.02% collagenase and 0.1 mM CaCl2 until the liver appeared soft. Next, the liver was removed and gently minced, and the released cells were dispersed in Dulbecco's modified Eagle medium (DMEM; Life Technologies) containing 10% FBS and 1% penicillin/streptomycin (Life Technologies). The solution containing the mixed cells and debris was passed through a 100-µm cell strainer (BD Bioscience, USA). Subsequently, the filtrate was centrifuged at 50 × g for 3 min at 4℃. The isolated cells were washed three times with DMEM and then seeded in collagen-coated plates. The cells were maintained in DMEM containing high glucose (4.5 g/L) supplemented with 10% FBS, 1% penicillin/streptomycin, 1 µg/mL insulin, and 10- 12 M dexamethasone for 24 h at 37℃ in a humidified atmosphere (5% CO2). The cells were then incubated with fresh Williams' E medium without FBS 24 h prior to the experiments.
Cell viability assay
Cell viability was measured using a CCK-8 assay. Mouse hepatocytes were cultured in 96-well plates (BD Bioscience) in triplicate for each group. The cells were treated with H2O2 with or without pretreatment with BHA or NAC for 30 min and cultured at 37℃ for 24 h. And 10 µL of CCK-8 solution was added to each well followed by further incubation at 37℃ for 3 h. Absorbance was measured at 450 nm using a microplate reader (BioTek Instruments, USA). All values are expressed as the mean ± standard error (SE) of triplicate experiments. The values were converted from absolute counts to a percentage of the control.
Western blot analysis
Cells were directly lysed in culture dishes with RIPA buffer (Boston BioProducts, USA) supplemented with proteinase and phosphatase inhibitor cocktail mixture (Thermo Scientific). Cell lysates (30 µg) were separated using 10% or 12% SDS-polyacrylamide gel electrophoresis and then transferred to polyvinylidene fluoride (PVDF) membranes (Millipore, Germany). The blots were then washed with Tris-buffered solution containing Tween-20 (TBST; 10 mM Tris-HCl [pH 7.6], 150 mM NaCl, and 0.1% Tween-20), blocked with 5% skimmed milk in TBST for 1 h at room temperature, and incubated for 12 h at 4℃ with the appropriate primary antibodies against Bax, Bcl-2, and PARP-1 at a 1:1,000 dilution. The membranes were then washed with TBST and incubated with horseradish peroxidase-conjugated goat anti-rabbit or goat anti mouse IgG antibodies (1:5,000 dilution) for 12 h at 4℃. The bands were visualized using an enhanced chemiluminescence detection system (Thermo Scientific) according to the manufacturer's protocols.
Detection of intracellular ROS
CM-H2DCFDA (DCF-DA), which functions as a ROSsensitive fluorophore, was used to detect intracellular ROS generation. Mouse hepatocytes on 35-mm cell culture dishes (BD Bioscience) were treated with H2O2 with or without pretreatment with BHA or NAC for 30 min and cultured at 37℃ for 2 h. The cells were moved into the dark and incubated with 5 µM DCF-DA for 30 min at 37℃. Next, the cells were washed three times with PBS and viewed using fluorescence microscopy (DM IL LED Fluo; Leica, Germany) at 100 × magnification. Fluorescence intensity was measured using Tecan Infinite M200 Pro (Life Technologies).
Results
BHA inhibits H2O2-induced apoptosis in primary cultured mouse hepatocytes
To investigate the effect of H2O2 on primary mouse hepatocytes, cell viability was measured with a CCK-8 assay. Fig. 1A shows that treatment of mouse hepatocytes with H2O2 at concentrations ranging from 100 to 1,000 µM for 24 h led to a significant decrease in cell viability in a dose-dependent manner. We also evaluated the expression of apoptotic marker proteins. H2O2 promoted Bax expression, decreased Bcl-2 levels, and increased PARP-1 cleavage. The Bax/Bcl-2 ratio also significantly increased in a dose-dependent manner.
To determine the effect of BHA (panel A in Fig. 2) on decreased cell viability caused by H2O2, cells were pretreated with various concentrations (0~10 µM) of BHA for 30 min and then treated with 1,000 µM H2O2. BHA prevented the decreases in cell viability caused by H2O2 (panel B in Fig. 2). These results indicate that BHA pretreatment protects primary cultured mouse hepatocytes from H2O2-induced apoptosis.
BHA ameliorates H2O2-induced oxidative stress in primary cultured mouse hepatocytes
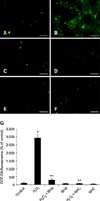
In order to determine whether the protective effects of BHA were associated with antioxidant activities in H2O2-treated mouse hepatocytes, intracellular ROS levels were measured. As shown in panels A-G of Fig. 3, H2O2 significantly increased intracellular ROS levels. This was significantly attenuated by pretreatment with BHA (5 µM) or NAC, a common ROS scavenger (1,000 µM). NAC was used as a positive control.
Protective effect of BHA against H2O2-induced apoptosis is mediated by antioxidant activity in primary cultured mouse hepatocytes
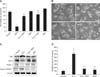
As shown in panel A of Fig. 4, decreased cell viability due to H2O2 was significantly inhibited by pretreatment with BHA or NAC. These results were confirmed by observing cell morphology (panel B in Fig. 4). Increased Bax levels, decreased Bcl-2 expression, and cleavage of PARP-1 promoted by H2O2 were attenuated by pretreatment with BHA (panel C in Fig. 4). Additionally, increases of the Bax/Bcl-2 ratio due to H2O2 treatment were also decreased by BHA (panel D in Fig. 4).
Discussion
Oxidative stress-induced cell damage has been implicated in various disorders. In particular, oxidative stress in the liver is related to apoptotic cell death that is associated with the development of liver disease [5]. H2O2 is a major ROS produced intracellularly during many physiological and pathological processes, and causes oxidative damage. This compound has been extensively used as an inducer of oxidative stress for in vitro models. Therefore, H2O2 was selected to promote oxidative damage in the current investigation.
It has been reported that BHA has an antioxidant effect possibly related to the attenuation of oxidative stress [11]. However, the mechanism underlying the hepatoprotective effect of BHA remains unclear. In the present study, it was demonstrated that BHA protected primary cultured mouse hepatocytes against H2O2-induced cytotoxicity by preventing oxidative stress and apoptosis.
Our results showed that H2O2 treatment significantly decreased cell viability while increasing Bax expression and reducing Bcl-2 levels. Members of the Bcl-2 family, which includes Bcl-2, Bcl-xL, Bad, and Bax, are important regulators of various apoptotic pathways [32]. It has been shown that Bax exerts pro-apoptotic effects whereas Bcl-2 possesses anti-apoptotic activity [37]. In general, Bcl-2 inhibits apoptosis by negatively regulating the apoptotic activity of Bax and forming Bcl-2/Bax heterodimers. The Bcl-2/Bax ratio can help determine whether a cell will live or die after being exposed to an apoptotic stimulus. In this study, a remarkable increase of the Bax/Bcl-2 ratio was observed after H2O2 treatment. Additionally, PARP-1 cleavage increased after H2O2 treatment. The cleavage of PARP is a hallmark of apoptosis [20]. PARP is known to be required for DNA repair. Pro-PARP is cleaved by activated apoptotic enzymes including caspase-3, a major mediator of apoptosis [6,13].
In this study, we demonstrated that H2O2 markedly increased cytotoxicity and intracellular ROS generation in mouse hepatocytes. Pretreatment with BHA significantly inhibited the decrease of cell viability as well as the increased intracellular ROS concentration. Additionally, our results showed that treatment with BHA down-regulates the H2O2-induced increase of the Bax/Bcl-2 ratio and PARP-1 cleavage. Previous studies have clearly shown that ROS induce apoptosis in hepatocytes [10,40]. ROS participate and regulate a diverse number of downstream signaling pathways that govern specific cellular functions [1,12] such as growth, metabolism, cell division, necrosis, apoptosis, and aging [7,18,24]. However, an imbalance in the formation and neutralization of ROS leads to oxidative stress [3]. Oxidative stress caused by ROS is responsible for a wide variety of cellular damage and is the most widely recognized mechanism of secondary injury [36]. Following oxidative stress, the overproduction of ROS and subsequent antioxidant depletion results in a total breakdown of the endogenous antioxidant defense mechanisms, culminating in the failure to protect cells from oxidative damage [16].
In conclusion, data from this study demonstrated that BHA possesses antioxidant activity. Additionally, pretreatment with BHA prevented apoptotic cell death induced by oxidative stress in mouse hepatocytes. Therefore, BHA may be beneficial for treating liver diseases caused by oxidative stress.


 XML Download
XML Download