

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Increased production and accumulation of melanin is a characteristic of many skin diseases, including acquired hyperpigmentation, melasma, postinflammatory melanoderma, and solar lentigo [835]. Melanin is one of the most widely distributed pigments, and is found in bacteria, fungi, plants, and animals. This compound is mostly produced in melanosomes of melanocytes that are dendritic cells found in the basal layer of the epidermis [16]. Abnormal accumulation of melanin in specific parts of the skin resulting in over-pigmented patches (e.g., melasma, freckles, ephelide, and senile lentigines) can be aesthetically undesirable [31]. Because acquired hyperpigmentation usually occurs in light-exposed areas of the skin, this condition has psychosocial and cosmetic relevance.
In human skin, melanin exists in two basic forms, black/brown eumelanin and yellow/red pheomelanin, that are synthesized from tyrosine via a common biosynthetic pathway [13]. The key enzyme in melanin biosynthesis is copper-dependent tyrosinase that catalyzes the initial rate-limiting step for conversion of tyrosine to 3,4-dihydroxyphenilalanine (DOPA), and is controlled by tyrosinase-related protein (TRP)-1 and TRP-2 [20]. Several depigmenting agents modulate skin pigmentation by regulating the transcription and activity of tyrosinase as well as that of TRP-1 and TRP-2. Tyrosinase inhibitors such as hydroquinone, kojic acid (KA), and arbutin (AT) have been used to treat hyperpigmention disorders but have recently been found to be unsafe for use in humans [101824]. Consequently, there is increasing interest in plant-derived extracts that can induce depigmentation without negative side effects on human health.
Tea is one of the most popular beverages worldwide. Fresh tea leaves contain polyphenols, primarily the catechins epigallocatechin (EGC), epicatechin (EC), epigallocatechin gallate (EGCG), and epicatechin gallate (ECG). These four catechins, of which EGCG is the most abundant, account for up to 30% of tea leaf dry weight [12]. Green tea (GT) is made from rolled and dried leaves that are steamed and/or pan-fried to inactivate endogenous polyphenol oxidase (PPO). White tea (WT) is unfermented tea produced from new growth buds and young leaves. The buds may be shielded from sunlight during growth to reduce chlorophyll contents, which makes the leaves appear white [2]. Black tea (BT) is produced without heat inactivation. The fermentation/oxidation phase after rolling enables large-scale PPO-catalyzed conversion of simple phenolics into more complex polyphenols such as thearubigins and theaflavins that are responsible for the characteristic BT reddish-brown color and astringency [32]. Most tyrosinase inhibitors are phenol/catechol derivatives, structurally similar to tyrosine or DOPA, and thus act as alternative tyrosinase substrates that are converted without pigment production [21]. Many studies have shown that phenolic compounds have antioxidant activity due to their capacity for scavenging free radicals and chelating metal ions along with an ability to prevent free radical formation [1262934].
In the present study, we investigated the anti-melanogenic effects of Camellia sinensis L. (family Theaceae) water extracts (CSWEs) and the potential mechanisms by which they induce skin depigmentation. The effects of CSWEs on tyrosinase activity, melanin synthesis, and expression of melanogenic enzymes at the protein and mRNA levels were evaluated in melan-A cells. These cells are immortalized mouse melanocytes characterized by high levels of tyrosinase and melanin expression [530].
Materials and Methods
Materials
Melan-A cells were obtained from Dr. Dorothy Bennett (St. George's Hospital, UK). EGC, EC, EGCG, ECG, a theaflavin standard mixture, AT, KA, 3-(4,5-dimethyl-thiazol-2-yl)-2, 5-diphenyl-tetrazolium bromide (MTT), 3,4-dihydroxy-L-phenylalanine (L-DOPA), L-tyrosine, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich (USA). TRP-1 and TRP-2 were purchased from Santa Cruz (USA). All reagents used in this study were of analytical grade.
Preparation of tea extracts
BT (China), GT (Korea), and WT (China) leaves were obtained from an oriental medicinal herb market (Korea). The leaves (600 g of each type) were boiled in 6 L of distilled water for 2 h in a heating extractor (COSMOS-660; Kyungseo Machine, Korea), concentrated, and powdered by lyophilization. Yields of the BT, GT, and WT extracts were 10.2, 21.3, and 20.0%, respectively.
Analysis of heavy metal contents
Tea extract samples (200 mg) were digested using 10 mL of concentrated HNO3 for 15 min in a microwave acid digestion system (MARS5; CEM, USA). The digests were diluted to 50 mL in a volumetric flask with deionized water. The total lead (Pb), arsenic (As), and cadmium (Cd) concentrations were then determined using an inductively coupled plasma mass spectrometer (ICP-MS; ELAN 9000; PerkinElmer, USA). Total mercury (Hg) levels were measured using 20 mg of CSWE in an automatic mercury analyzer (MA-2; Nippon Instruments Corporation, Japan).
Measurement of catechin and theaflavin contents
Catechin levels were measured with an HPLC-photodiode array detector (Waters 2996 PDA Detector; Waters, USA) and a liquid chromatography system (Waters e2695; Waters). The extract sample (10 µL) was injected into the HPLC column (5 µm, 250 × 4.6 mm, Shiseido CAPCELL PAK C18 UG 120; Shiseido, Japan). Mobile phase A was composed of 0.1% acetic acid and mobile phase B was composed of 100% acetonitrile. The catechins were eluted with 95% mobile phase A at 0 min, 75% mobile phase A at 20 min, 100% mobile phase B at 21 min, and 95% mobile phase A at 36 min. The flow rate was 1.0 mL/min at 40℃. Peaks were monitored at 280 nm and UV spectra were recorded.
Theaflavins were quantified with an HPLC-photodiode array detector (Waters 2998 PDA Detector; Waters) and a liquid chromatography system (Waters e2695; Waters). The extract sample (10 µL) was injected into the HPLC column as described above. Mobile phase A was composed of 0.1% acetic acid:acetonitrile:tetrahydrofuran (96:2:2), and mobile phase B was 100% acetonitrile. The theaflavins were eluted with 100% mobile phase A at 0 min, 40% mobile phase A at 45 min, and 100% mobile phase A at 47 min. The flow rate was 1.0 mL/min at 40℃. Peaks were monitored at 310 nm and UV spectra were recorded. The individual catechins and theaflavins were identified by comparing the retention times of the analytes with those of reference standards.
Analysis of antioxidant contents
Total polyphenol contents of the tea extracts were evaluated using a Folin-Denis assay [11]. Total flavonoid contents were assessed as previously described by Davies et al. [9] with modifications. The test material (100 µL) was transferred into test tubes before 1 mL of di (ethylene glycol) reagent and 100 µL of 1 N NaOH were added. The mixture was shaken vigorously and incubated in hot water at 37℃ for 60 min. Absorbance was then measured at 420 nm.
Cell culturing
Melan-A cells were grown in Roswell Park Memorial Institute (RPMI)-1640 medium supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin (P/S), and 200 nM 12-O-tetradecanoylphorbol-13-acetate (TPA) at 37℃ in a 10% CO2 incubator for 72 h prior to the experiment.
Cell viability assay
Cell viability was assessed with an MTT assay. Melan-A cells were seeded in a 96-well plate (0.5 × 104 cells/well) for 24 h and then treated with 200 µL of the CSWEs (0~50 µg/mL) in RPMI-1640 medium for 48 h. The medium was then replaced with RPMI-1640 medium containing 0.5 µg/mL MTT and the cells were further incubated for 3 h. After centrifugation at 168 × g for 10 min to pellet the cells, the medium was removed, 200 µL of DMSO were added, and the solution was incubated for 15 min with shaking to resuspend the cells. Cell viability was assessed by measuring absorbance at 540 nm with an ELISA reader (680; Bio-Rad Laboratories, Japan).
Melanin assay and morphological evaluation
Melan-A cells were seeded in a 48-well plate (2 × 104 cells/well) for 24 h and then treated with 500 µL of CSWEs (0~12.5 µg/mL) in RPMI-1640 medium for 72 h. Thereafter, the cells were washed and incubated with fresh tea extract solutions for another 72 h. The cells were observed with a model CKX41SF microscope (Olympus, Japan) and photographed at 200× magnification using a U-TVO.5XC-3 digital camera (Olympus) equipped with DMC e310 software (INS Industry, Korea). Next, the cells were dissolved in 1 N NaOH, and absorbance was measured at 490 nm (OD 490) using an ELISA reader. Melanin content was estimated as the OD 490 value/µg of protein and expressed as a percentage relative to the untreated control value (100%).
Tyrosinase activity assay
For the intracellular tyrosinase activity assay, melan-A cells were seeded in 60-mm cell culture dishes (4 × 105 cells/well) for 24 h and then treated with 5 mL of tea extracts (0~12.5 µg/mL) for 48 h. The cells were washed with phosphate buffered saline, detached with 200 µL of 1% Triton X-100, transferred to Eppendorf tubes, subjected to extraction on ice with agitation six times every 10 min, and centrifuged at 18,620 × g for 20 min at 4℃. Next, 100 µL of L-DOPA were added and the mixture was incubated at 37℃ with 10% CO2 for 1 h. The OD 490 nm was then measured using an ELISA reader. Tyrosinase activity was estimated as the OD 490/µg protein/min and expressed as a percentage of the untreated control value (100%). For tyrosinase activity in the cell extract, the cultured melan-A cells were centrifuged, and 50 µL of supernatant was mixed with 49 µL of 0.1 M phosphate buffer (pH 6.8) and 1 µL of tea extracts (0~12.5 µg/mL). Subsequently, 100 µL of 0.2% L-DOPA was added, the absorbance was measured, and the percentage activation was calculated.
Western blotting
Melan-A cells treated with the CSWEs (6.25, 12.5, and 25.0 µg/mL) for 48 h were lysed by sonication in 0.1 M Tris-HCl (pH 7.2) buffer containing 1% Nonidet P-40, 0.01% SDS, and protease inhibitor cocktail (Roche, Germany). Protein concentration in the cell lysates was measured using a protein assay kit (Pierce, USA) with bovine serum albumin as the standard. Equal amounts of protein (10 µg) were separated by electrophoresis in a 10% polyacrylamide gel, transferred onto nitrocellulose membranes, and incubated with antibodies against tyrosinase/prolyl endoprotease-7 (PEP-7, 1:10,000 dilution), TRP-1/PEP-1 (1:10,000), and TRP-2/PEP-8 (1:10,000) kindly provided by Dr. Vincent J. Hearing (National Institutes of Health [NIH], USA). The membranes were then incubated with horseradish peroxidase-conjugated anti-rabbit IgG (1:1,000; Amersham, UK). Antibody binding was detected by chemiluminescence using electrochemical luminescence (ECL) reagents (Amersham). Band intensity was semi-quantified using the Image J program (NIH); β-actin was used as an internal control.
Reverse transcription (RT)-PCR
Total RNA was isolated from cells treated with 6.25 and 12.5 µg/mL tea extracts for 48 h using TRIzol reagent (Life Technologies, USA) according to the manufacturer's instructions. Equal amounts of total RNA (5 µg) were reverse transcribed in a 40-µL reaction mixture containing 8 µL of Molony murine leukemia virus reverse transcriptase (M-MLV RT), 5× buffer, 3 µL of 10 mM dNTPs, 0.45 µL of 40 U/µL RNase inhibitor, 0.3 µL of 250 U/µL M-MLV RT (Promega, USA), and 3.75 µL of 20 µM oligo dT (Bioneer, Korea). Single-stranded cDNA was then used as a template for PCR amplification in a reaction containing 4 µL of 5× green Go Taq Flexi buffer, 0.4 µL of 10 mM dNTPs, 0.1 µL of 5 U/µL Taq polymerase, 1.2 µL of 25 mM MgCl2 (Promega), and 0.4 µL of each primer (20 µM). The primer sequences were as follows: 5'-CAT TTT TGA TTT GAG TGT CT-3' (forward), 5'-TGT GGT AGT CGT CTT TGT CC-3' (reverse) for tyrosinase; 5'-GCT GCA GGA GCC TTC TTT CTC-3' (forward), 5'-AAG ACG CTG CAC TGC TGG TCT-3' (reverse) for TRP-1; 5'-GGA TGA CCG TGA GCA ATG GCC-3' (forward), 5'-CGG TTG TGA CCA ATG GGT GCC-3' (reverse) for TRR-2; and 5'-ACC GTG AAA AGA TGA CCC AG-3' (forward), 5'-TAC GGA TGT CAA CGT CAC AC-3' (reverse) for β-actin. PCR conditions for tyrosinase and TRP-1 were as follows: 28 cycles of denaturation at 94℃ for 60 sec, annealing at 56℃ for 60 sec, and extension at 72℃ for 60 sec. PCR conditions for TRP-2 were 28 cycles of denaturation at 94℃ for 60 sec, annealing at 64℃ for 60 sec, and extension at 72℃ for 60 sec. PCR for β-actin was carried out with 30 cycles of denaturation at 94℃ for 30 sec, annealing at 51℃ for 30 sec, and extension at 72℃ for 60 sec. The PCR products were separated on 1.2% agarose gels. Expected sizes of the amplified tyrosinase, TRP-1, TRP-2, and β-actin fragments were 1192, 268, 1044, and 528 bp, respectively. DNA band density was semi-quantitatively analyzed using a Kodak Gel Logic 100 image analysis system (Eastman Kodak, USA). The gene expression of tyrosinase, TRP-1, and TRP-2 was normalized relative to β-actin as an internal control.
Results
Effects of the tea extracts on melan-A cell viability
To examine the safety of the CSWEs as an herbal medicine, we evaluated toxicity of the extracts by analyzing their heavy metal contents. Pb, As, Hg, and Cd levels in BT, GT, and WT were below the maximum permissible levels for herbal medicines set by the Ministry of Food and Drug Safety (MFDS). The Hg concentration in all three extracts was below the analytical detection limit (Table 1). These results indicated that the tea extracts could be safely used as herbal medicines and in cosmetics.
To further examine the potential toxicity of BT, GT, and WT, the viability of melan-A cells exposed to the extracts at concentrations between 0 and 50 µg/mL was measured for 48 h. The well-known depigmenting agents AT and KA along with the primary tea catecholamine EGCG were included as positive controls. Cells were viable in the presence of all concentrations of AT, KA, and BT similar to the untreated controls (Fig. 1). At 25 µg/mL, GT and WT also increased cell viability; however, a higher concentration (50 µg/mL) reduced melan-A cell viability to 62% and 76% relative to the untreated control, respectively. EGCG was found to have the greatest cytotoxicity among the tested compounds. At 25 and 50 µg/mL, this reagent decreased cell viability to 38% and 6% relative to the control group, respectively. Thus, BT, KA, and AT had the weakest cytotoxicity (≤ 50 µg/mL), followed by GT and WT (≤ 25 µg/mL), and EGCG (≤ 12.5 µg/mL).
Polyphenol and flavonoid contents of the tea extracts
The total polyphenol concentrations of AT, KA, EGCG, BT, GT, and WT were 24, 32, 435, 117, 112, and 123 mg/g, respectively, as determined using the tannic acid standard curve (Table 2). The total flavonoid concentrations of AT, KA, EGCG, BT, GT, and WT were 13, 14, 180, 82, 77, and 85 mg/g, respectively, according to the rutin standard curve (Table 2). The tea extracts were found to contain much higher levels of total polyphenols and flavonoids than AT and KA. Among the teas, WT had the highest concentrations followed by BT and GT.
Catechin and theaflavin contents of the tea extracts
GT (196.58 ± 0.81 mg/g) contained greater amounts of catechins than WT (137.48 ± 0.79 mg/g) and BT (15.30 ± 0.60 mg/g). In contrast, BT contained 1.49 ± 0.10 mg/g of theaflavins including theaflavin, theaflavin-3 gallate, theaflavin-3' gallate, and theaflavin-3,3' digallate, which were not detected in GT or WT (Table 3).
Effects of the tea extracts on melanin synthesis
To assess the hypopigmentation effect of the tea extracts, melan-A cells were exposed to the test reagents at concentrations ranging from 0 to 12.5 µg/mL for 72 h and examined by microscopy. The untreated control cells contained abundant black precipitates indicative of melanin synthesis and well-developed dendrites (panel A in Fig. 2). On the other hand, melanin deposition and dendrite formation were conspicuously reduced in the cells treated with BT, GT, and WT.
We also quantitatively analyzed melanin contents of the cells. AT, KA, EGCG, BT, GT, and WT reduced melanin synthesis compared to the control in a concentration-dependent manner (data not shown). At a dose of 12.5 µg/mL, AT, KA, EGCG, BT, GT, and WT decreased melanin contents by 18% (p < 0.00l), 7% (p < 0.05), 29% (p < 0.00l), 52% (p < 0.00l), 20% (p < 0.00l), and 31% (p < 0.00l) compared to that observed in untreated cells, respectively (panel B in Fig. 2). The tea extracts were found to more significantly inhibit melanin synthesis in melan-A cells than AT and KA. Among the teas, BT showed the highest inhibitory effect which was superior even compared to that of EGCG.
Effects of the tea extracts on tyrosinase activity
Tyrosinase is the rate-limiting enzyme in melanin synthesis. To examine the inhibitory effect of the tea extracts on tyrosinase activity, melan-A cells were exposed to the test materials at concentrations between 0 and 12.5 µg/mL for 48 h. AT was used as a positive control because this compound was found to exert a greater inhibitory effect on melanin synthesis than KA. Compared to the untreated control group, tyrosinase activity in cells treated with AT, EGCG, BT, GT, and WT was reduced in a concentration-dependent manner (data not shown). At 12.5 µg/mL, intracellular tyrosinase activity was decreased by 12%, 61%, 52%, 45%, and 58%, respectively (p < 0.001; Fig. 3), indicating that tyrosinase activity in mealan-a cells was reduced more by the tea extracts than AT. Among the teas, WT had the greatest inhibitory effect, which was similar to that of EGCG. Additionally, treatment with 12.5 µg/mL of AT, EGCG, BT, GT, and WT decreased cell-extracted tyrosinase activity by 13%, 18%, 25%, 15%, and 16%, respectively (p < 0.001; data not shown). Among the extracts, BT had the most potent inhibitory effect, superior even to that of EGCG. These results suggest that the inhibition of melanin synthesis by CSWEs is related to decreased melanocyte tyrosinase activity.
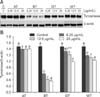
Effects of the tea extracts on tyrosinase protein and mRNA expression
To determine whether the inhibition of tyrosinase activity by CSWEs was associated with tyrosinase expression, melan-A cells were exposed to the extracts at concentrations ranging from 0 to 25 µg/mL for 48 h. Tyrosinase protein expression was then measured by Western blotting (panel A in Fig. 4). Treatment with BT, GT, and WT reduced tyrosinase protein expression in a concentration-dependent manner (panel B in Fig. 4). At 12.5 µg/mL, BT, GT, and WT significantly (p < 0.001) decreased tyrosinase protein levels by 72%, 18%, and 25%, respectively, compared to the control. In contrast, AT at all tested concentrations did not affect the protein expression of tyrosinase. We also measured the protein expression of the melanogenic enzymes TRP-1 and TRP-2, and found that the levels of both proteins were not altered by AT or CSWE treatment (data not shown).
We also performed RT-PCR to examine the mRNA levels of tyrosinase, TRP-1, and TRP-2 in cells exposed to the extracts at concentrations ranging from 0 to 12.5 µg/mL for 48 h (panel A in Fig. 5). No difference in mRNA expression of melanogenic enzymes was observed between the treated and control cells (panel B in Fig. 5). These results suggest that the inhibitory effect of tea extracts on cellular tyrosinase activity is related to decreased tyrosinase protein expression.
Discussion
Tea is one of the most popular beverages in the world and is also known for its health-promoting effects. Four types of tea are produced from the leaves of Camellia sinensis: WT, GT (both unfermented teas), oolong tea (semi-fermented tea), and BT (fermented tea). In a previous study, oolong tea water extract was reported to inhibit melanogenesis in B16 mouse melanoma cells [4]. However, the melanogenic effects of BT, GT, and WT have not been sufficiently investigated. In the present study, we examined the hypopigmentation effects of BT, GT, and WT (CSWEs) in immortalized melanocytes.
EGCG, a predominant tea catechin, has been reported to more effectively inhibit melanin synthesis than KA in the spontaneously immortalized mouse melanocyte cell line Mel-Ab [15]. Additionally, oolong tea water extract was found to exert a greater inhibitory effect on melanogenesis in B16 melanoma cells than ascorbic acid [4]. In the current study, we discovered that BT and WT had higher levels of anti-melanogenic activity than AT, KA, and EGCG. In addition, BT more effectively reduced melanin deposition and dendrite formation along with total melanin contents than GT and WT. HPLC analysis revealed that BT contained theaflavins (1.49 ± 0.10 mg/g) that were not detected in GT or WT. Theaflavins, a mixture of theaflavin, theaflavin-3-gallate, theaflavin-3'-gallate, and theaflavin-3,3'-digallate, are formed by two catechin monomers via the action of PPO during fermentation. Among these compounds, theaflavin-3,3'-digallate was previously found to prevent melanogenesis more efficiently than EC, ECG, EGC, EGCG, and KA in mouse B16 melanoma cells while the other theaflavins had no significant anti-melanogenic effect [36]. Therefore, it is believed that the depigmentation effects of CSWEs can be augmented by fermentation, and the potential depigmentation component of BT might be theaflavin-3, 3'-digallate.
Tyrosinase, a rate-limiting enzyme in melanogenesis, is a melanocyte-specific copper-containing glycoprotein located within specialized organelles called melanosomes [20]. Tea and its components have been shown to inhibit intracellular and cell-extracted tyrosinase activities. EGCG causes such inhibition in mushrooms [1523], and oolong tea reduces tyrosinase activity in B16 mouse melanoma cells [4]. Tea flavonoids have metal chelation abilities [26]. For example, proanthocyanidins, which are flavonoids from grape seeds, inhibit tyrosinase by chelating tyrosinase-bound copper [34]. BT theaflavins and GT flavonols are also known to exert antioxidant effects by radical scavenging and metal chelation [28]. WT was also reported to have higher levels of antioxidant activity than GT [33].
In the present study, we found that the levels of flavonoids in EGCG, BT, GT, and WT were 180, 82, 77, and 85 mg/g, respectively, which may account for the inhibition of tyrosinase activity. Among the teas, WT exerted the greatest inhibitory effect on intracellular tyrosinase activity followed by BT and GT. In contrast, BT had the highest inhibitory effect on cell-extracted tyrosinase activity followed by WT and GT. Of these two types of tyrosinase activities, cell-extracted one was coincident with melanin synthetic activities of CSWEs in the current study. Although tyrosinase may be inhibited through different mechanisms (e.g., blocking of catalytic or regulatory sites as well as protein degradation) [25] that remain unclear in melan-A cells, our results suggest that CSWEs may inhibit tyrosinase activity via copper chelation. Recently, rhododendrol [4-(4-hydroxyphenyl)-2-butanol], a depigmentation-inducing phenolic compound developed as a lightening/whitening cosmetic by Kanebo Cosmetics Inc. (Japan) was reported to cause depigmentary disorders such as leukoderma and idiopathic vitiligo [1427]. This reagent was also found to be cytotoxic against human melanocytes at high concentrations, suggesting that tyrosinase-dependent accumulation of endoplasmic reticulum stress and/or activation of the apoptotic pathway may contribute to melanocyte cytotoxicity. Although BT had the lowest cytotoxicity among the tested reagents, and WT and GT had lower cytotoxicity than EGCG in the current investigation, further studies should be conducted to evaluate depigmentary disorders associated with CSWEs. Additionally, the relationship between cytotoxicity and molecular events following CSWE-tyrosinase interaction should be clarified.
CSWEs, especially BT, also significantly (p < 0.001) decreased tyrosinase protein expression in a concentration-dependent manner. In contrast, AT did not affect tyrosinase protein levels. This finding is consistent with ones from previous studies showing that AT does not change the expression of tyrosinase, TRP-1, or TRP-2 in B16 melanoma cells [17], melan-A cells [37], and human melanocytes [6]. These data indicate that AT depigmentation activity may be based on post-translational inhibition of tyrosinase activity [619]. It has been reported that oolong tea decreases tyrosinase protein and mRNA levels in B16 mouse melanoma cells, thereby inhibiting melanogenesis at the transcriptional level [4], whereas EGCG reduces tyrosinase expression in Mel-Ab cells at the protein level [15]. In the present study, we found that CSWEs decreased tyrosinase protein expression but did not affect mRNA levels in melan-A cells, suggesting that the teas may decrease the post-transcriptional stability of tyrosinase and accelerate its degradation [3].
Our results are in agreement with those from a previous report showing that danazol inhibits melanogenesis in B16 mouse melanoma cells by downregulating the expression of tyrosinase protein but not mRNA [7]. In addition, ethanol extracts of traditional Chinese herbs (Radix ophiopogonis and Folium perillae) also cause depigmentation in melan-A cells by suppressing tyrosinase activity at the post-transcriptional level [37]. Overall, these results indicate that herbal extracts may have diverse mechanisms of depigmentation due to their complex composition.
In conclusion, our study has demonstrated that CSWEs (BT, GT, and WT) inhibit melanogenesis more efficiently than AT in melan-A cells. Among the different teas, BT (fermented tea) may be the most effective depigmentation agent with the lowest cytotoxicity and highest anti-melanogenic activities that are superior to those of EGCG. The potential mechanism underlying the depigmentation effects of CSWEs is the inhibition of tyrosinase activity. This is due to accelerated tyrosinase degradation and reduced levels of the protein, but not modulation of mRNA transcription. The potential depigmentation reagents in BT are theaflavins. However, this hypothesis should be further investigated by isolating and identifying biologically active components contained in BT.







 XML Download
XML Download