

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Melatonin is an indolamine derived from tryptophan. Biosynthesis of this factor is mediated by the sequential reaction of two key enzymes: arylalkylamine N-acetyltransferase (AANAT) and hydroxyindole-O-methyltransferase (HIOMT) [8]. AANAT catalyzes the transfer of an acetyl group from acetyl-CoA to the primary amine of serotonin that leads to the production of N-acetylserotonin (NAS) [11,31]. Many studies have demonstrated that melatonin has chronobiotic, antioxidant, anti-inflammatory, neuroprotective, and analgesic properties [3,4,6,7,26]. Melatonin exerts theses physiological functions via its G protein-coupled receptors: MT1 and MT2 [5].
AANAT and MT2 mRNA expression has been found in the central nervous system including the hippocampus, cerebellum, and spinal cord [23,30]. The effects of melatonin on the brain have been well established. However, the possibility of direct melatonin action in the spinal cord has not been extensively investigated. Recently, studies have suggested that melatonin influences antinociceptive effects via MT2 in the spinal cord [28,30].
Melatonin secretion progressively declines during the normal aging process. A significant reduction of circulating melatonin has also been observed with neurodegenerative disorders such as Alzheimer's disease [10,12]. However, few studies monitoring AANAT and MT2 expression in the spinal cord, which is morphologically, neurochemically, and physiologically altered during normal aging, have been performed [19,27,29]. Therefore, we conducted the present study to compare patterns of AANAT and MT2 immunoreactivity in the spinal cord of young adult and aged beagle dogs, which is a good model of aging [2,20].
Materials and Methods
Experimental animals
Clinically and neurologically normal male beagle dogs were obtained from the Experimental Animal Center, Kangwon National University (Korea). The animals were used when they were 2~3 years old (young adult dogs, n = 12, 9~11 kg body weight) or 10~12 years old (aged dogs, n = 12, 10~13 kg body weight). None of the dogs showed any clinical symptoms or other signs of neural disorders. The animals were housed in a conventional environment with adequate temperature (23℃) and humidity (60%) as well as a 12-h light/dark cycle with light onset at 07:00 am. They also had free access to water and food (DBL, Korea). To ensure adaptation to the new environment, the animals were kept in the departmental holding room for 1 week prior to the experiments. Procedures for handling and caring for the dogs were in compliance with current international laws and policies (National Institutes of Health [NIH] Guide for the Care and Use of Laboratory Animals, NIH Publication No. 85 23, 1985, revised 1996) [16].
Tissue processing for histology
Spinal cord tissue sections were prepared for histochemical analysis using a previously described method [1]. In brief, the young adult and aged dogs (n = 7 for each group) were anesthetized with a mixture of Zoletil 50 (8 mg/kg, Virbac Korea, Korea) and Xylazine (2 mg/kg, Bayer Korea, Korea), and perfused transcardially with 0.1 M phosphate-buffered saline (PBS, pH 7.4) followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB, pH 7.4). The cervical (C6-C8) and lumbar (L5-L6) spinal cord, levels that are closely related to limb innervation, were removed and postfixed in the 4% paraformaldehyde for 12 h at 4℃. The spinal cord tissues were cryoprotected by infusion with 30% sucrose overnight at 4℃. Next, the frozen tissues (-15℃~-20℃) were serially cut with a cryostat (Leica Biosystems, Germany) into sections 30 µm in thickness and transferred to six-well plates (SPL Lifescience, Korea) containing PBS.
Immunohistochemistry
Immunohistochemistry specific for AANAT and MT2 was performed under the same conditions for the dogs of different ages in order to determine whether the degree of staining was accurate. The sections were stained using a previously described method [1]. In brief, the sections were sequentially treated with 0.3% H2O2 and 10% normal goat serum (Vector Laboratories, USA) at room temperature. They were then incubated with rabbit anti-AANAT (diluted 1 : 500, Catalog No. ABIN751063; antibodies-online, USA) and goat anti-MEL-1B-R (T-18) (diluted 1 : 200, Catalog No. sc-13177; Santa Cruz Biotechnology, USA) overnight at 4℃. Next, the sections were subsequently exposed to biotinylated goat anti-rabbit IgG or rabbit anti-goat for 2 h and streptavidin peroxidase complex (1 : 200, Vector Laboratories) for 1 h at room temperature. Antibody binding was visualized by staining with 0.05% 3,3'-diaminobenzidine tetrahydrochloride (Sigma, USA) in 0.1 M Tris-HCl buffer (pH 7.2), and the sections were dehydrated by ethanol before being mounted on gelatin-coated slides (Marienfeld Superior, Germany) in Canada balsam (Kanto Chemical, Japan). In order to assess the immunostaining specificity, a negative control test was carried out with 10% pre-immune serum instead of primary antibody. No immunoreactivity was observed in any structure in the negative control.
Double immunofluorescence
To confirm the cellular location of AANAT immunoreactivity, sections of aged dog spinal cord were subjected to double immunofluorescence staining. Staining was performed using rabbit anti-AANAT (diluted 1 : 200, Catalog No. ABIN751063; antibodies-online, Germany) and mouse anti-NeuN (diluted 1 : 500, Catalog No. MAB377; EMD Millipore, USA) antibodies. The sections were incubated in the mixture overnight at room temperature. After washing three times for 10 min with PBS, the sections were then incubated in a mixture of Cy3-conjugated goat anti-rabbit IgG (1 : 200; Jackson ImmunoResearch Laboratories, USA) and fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG (1 : 200; Jackson ImmunoResearch Laboratories) for 2 h at room temperature. Immunoreactivity was observed with confocal microscopy (LSM 510 META NLO; Carl Zeiss, Germany).
Western blot analysis
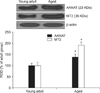
To confirm changes in AANAT and MT2 expression in the lumbar spinal cord of young adult and aged dogs, Western blot analysis was performed using our previously described method [14]. In brief, the spinal cord tissues were homogenized (Sigma) in 50 mM PBS (pH 7.4, Sigma-Aldrich, USA) containing EGTA (pH 8.0, Santa Cruz Biotechnology), 0.2% NP-40 (Sigma), 10 mM EDTA (pH 8.0, Sigma), 15 mM sodium pyrophosphate (Santa Cruz Biotechnology), 100 mM β-glycerophosphate (Santa Cruz Biotechnology), 50 mM NaF (Santa Cruz Biotechnology), 150 mM NaCl (Santa Cruz Biotechnology), 2 mM sodium orthovanadate (Santa Cruz Biotechnology), 1 mM phenylmethylsulfonyl fluoride (PMSF, Santa Cruz Biotechnology), and 1 mM dithiothreitol (DTT, Santa Cruz Biotechnology). After centrifugation at 16,000 × g in a microcentrifuge for 5 min, the protein concentration in the supernatants was determined using a Micro BCA protein assay kit (Pierce Biotechnology, USA). Aliquots of the supernatant containing 20 µg of total protein were boiled in loading buffer containing 150 mM Tris (pH 6.8, Santa Cruz Biotechnology), 3 mM DTT (Santa Cruz Biotechnology), 6% SDS (Santa Cruz Biotechnology), 0.3% bromophenol blue (Santa Cruz Biotechnology), and 30% glycerol (Junsei Chemical, Japan). The samples (20 µg of total protein) were then loaded onto a polyacrylamide gel. After electrophoresis, the separated proteins were transferred to nitrocellulose membranes (Pall Corporation, USA). The membranes were incubated with 5% dry non-fat milk (Santa Cruz Biotechnology) in PBS containing 0.1% Tween 20 (Santa Cruz Biotechnology) followed by incubation with rabbit anti-AANAT (diluted 1 : 1,000) and goat anti-MEL-1B-R(T-18) (diluted 1 :1,000), peroxidase-conjugated donkey anti-rabbit IgG (Sigma), or anti-goat IgG (Sigma). Antibody binding was detected with an enhanced luminol-based chemiluminescent (ECL) kit (Thermo Scientific, USA).
Data analysis
For quantitative analysis of AANAT and MT2 immunoreactivity, 10 sections per animal were randomly selected from the corresponding areas of the cervical (C6-C8) and lumbar (L5-L6) spinal cord according to our previously described method [1]. The images were converted into an array of 512 × 512 pixels corresponding to a tissue area of 500 × 500 µm (20× primary magnification). Data are expressed as a percent relative to that of the adult dogs. Images of AANAT- and MT2-immunoreactive structures were obtained with an AxioM1 light microscope (Carl Zeiss) equipped with a digital camera (AxioCam; Carl Zeiss) connected to a PC monitor (Samsung, Korea). Each pixel resolution was 256 gray levels. The staining intensity of all AANAT- and MT2-immunoreactive structures was evaluated based on the relative optical density (ROD) obtained after transformation of the mean gray level using the formula: ROD = log (256/mean gray level). The ROD of the complete field was measured. Next, the brightness and contrast of each image file were calibrated using Adobe Photoshop (ver. 8.0; Microsoft, USA), and then analyzed using NIH Image 1.59 software (developed at the NIH, USA). Background staining values were obtained and subtracted from the immunoreactive intensities. The Western blot was densitometrically scanned (Samsung) to quantify the ROD of each band using Scion Image software (Informer Technologies, USA). A ratio of the ROD was calculated as a percentage with the adult group value designated as 100%.
Results
AANAT immunoreactivity patterns and protein expression
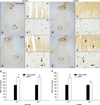
AANAT-immunoreactive cells in the adult dogs were distributed in the gray matter of the cervical and lumbar spinal cord (Figs. 1A and D), and AANAT-specific signals were generally found in the nuclei of spinal neurons (Figs. 1a, b, e, and f). In the aged dogs, the AANAT expression pattern was similar to that found in the young adult dogs (Figs. 1B and E). However, AANAT immunoreactivity in the nuclei of the spinal neurons was significantly increased in the aged dog spinal cord compared to that in the young adult spinal cord (Figs. 1c, d, C, g, h, and F). Western blotting showed that the AANAT protein level in the spinal cord of the aged group was markedly increased compared to that found in the young adult group (Fig. 2).
Double immunofluorescence staining for AANAT and NeuN
We performed double immunofluorescence staining specific for AANAT and NeuN (a neuron marker) to determine whether AANAT immunoreactivity co-localized with NeuN-positive spinal neuron nuclei. AANAT-positive structures were co-localized with NeuN-positive spinal neuron nuclei in the aged spinal cord (Fig. 3).
MT2-specific immunoreactivity and protein expression
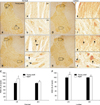
In the young adult dogs, MT2-immunoreactive cells were easily observed throughout the gray matter of the cervical and lumbar spinal cord. Additionally, moderate levels of MT2 immunoreactivity were found in the cytoplasm of spinal neurons (Figs. 4a, b, e, and f). In the aged dogs, the expression pattern of MT2 was similar to that appearing in the young adult dogs (Figs. 4B and E). However, the level of MT2-specific immunoreactivity was much higher in the aged dog spinal cord than in the young adult spinal cord (Figs. 4c, d, C, g, h, and F). In addition, strong MT2 immunoreactivity was detected in the cytoplasm of spinal neurons in the aged group (Figs. 4d and h). Results of the Western blot study demonstrated that MT2 protein levels in the spinal cord of the aged group were markedly increased compared to that in the adult group (Fig. 2).
Discussion
In the present study, we investigated age-related changes in the expression of AANAT and MT2 in the cervical and lumbar spinal cord. Levels of these factors are closely related with the innervations of limbs. Comparisons were made between young adult and aged dogs.
We found that AANAT-specific immunoreactivity generally appeared in nuclei of spinal neurons in the gray matter of the young adult spinal cord. This result concurs with data from a previous study showing that AANAT-immunoreactive structures can be observed in the nuclei of pinealocytes in humans and sheep [15]. In addition, it was reported that AANAT mRNA is mostly found in the gray matter of rat spinal cord [22,23]. It is thus likely that AANAT expression has a physiological function in spinal neurons.
The distribution pattern of MT2-immunoreactive cells in the spinal cord appears different among some species. It has been reported that melatonin receptors in rabbit and chicken spinal cord are localized in the dorsal horn and central gray matter [24,25]. Conversely, MT2 transcripts in the rat spinal cord are present in the dorsal and ventral horn [30]. Similar to rat spinal cord, MT2 immunoreactivity in the present study was observed in the gray matter of the cervical and lumbar spinal cord. The discrepancy may be species-related and reflect different functions of melatonin depending on the physiologically relevant location of the melatonin receptors.
It is well known that melatonin levels can change in a day and throughout the year [10,12,18]. This fluctuation in melatonin levels can be affected by a key enzyme, AANAT. In the present study, AANAT-specific immunoreactivity was significantly increased in the aged dog spinal cord compared to that in the young adult dogs. Our finding corresponds to data from a study by Sanchez-Hidalgo et al. [19] who reported that AANAT enzymatic activity was increased in the liver of aged rats although melatonin levels decrease during the aging process. In addition, age-related increases of AANAT mRNA levels and AANAT enzymatic activity have been found in the chicken pineal gland during postembryonic development [17].
In the current study, MT2-specific immunoreactivity was significantly increased in the aged dog spinal cord compared to that found in the young adult dogs. The level of immunoreactivity was increased in the neuronal cytoplasm. This result concurs with findings from a previous study showing that MT2 immunoreactivity and protein expression are markedly increased in the aged gerbil hippocampus [13]. Furthermore, it was discovered that plasma melatonin concentration are lowest although the affinity of melatonin for 2-[125I]-iodomelatonin and total 2-[125I]-iodomelatonin along with melatonin receptor mRNA expression are highest in the pars tuberalis of rat and in the pars tuberalis and suprachiasmatic nuclei of hamster [9,21].
In summary, we found that AANAT and MT2 immunoreactivity and protein levels increased in the nuclei of spinal neurons from aged dog spinal cord. These data suggest that increased AANAT and MT2 immunoreactivity in aged spinal cord neurons might be associated with a feedback mechanism that compensates for decreased melatonin production during the normal aging process. Based on the results of this study, a further study on the physiological function of melatonin in spinal neurons needs to find more specific mechanisms of aging-related changes in AANAT and MT2.

 XML Download
XML Download