

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Clostridium
(C.) difficile is an obligate, anaerobic, Gram-positive, spore-forming bacillus that is the most common infectious cause of antibiotic-associated colitis. Although the organism usually exists in a vegetative form that is highly sensitive to oxygen, it can also exist as a spore which is heat-stable and able to survive a variety of harsh conditions. The use of broad-spectrum antibiotics to treat diseases can disturb the normal bacterial flora in the intestine, resulting in host susceptibility to colonization and overgrowth of toxigenic C. difficile that can cause C. difficile infection (CDI). CDI produces a wide spectrum of clinical symptoms in humans ranging from mild diarrhea to severe life-threatening colonic perforation and toxic megacolon [4]. A very similar scenario of severe and often fatal antibiotic-associated diarrhea has been reported in adult horses [32]. Recently, researchers also noticed that C. difficile-associated diarrhea outbreaks tend to occur on breeding farms, particularly among foals during their first week of life [5].
C. difficile produces a number of virulence factors that contribute to its pathogenicity. However, the main virulence factors are two large clostridial toxins A (TcdA) and B (TcdB). Over 400 different types of C. difficile have been identified by HindIII restriction enzyme analysis [1]. Among these, only toxin-producing strains cause disease. Although the virulence of TcdB is well recognized, both toxins are important for causing disease [21]. Both compounds are exotoxins that can bind to intestinal epithelial cells and induce inflammation, fluid and mucous secretion, and damage to the intestinal mucosa [21]. TcdA and TcdB belong to a family of large clostridial toxins that share certain common structural and functional domains [1].
Vancomycin and metronidazole have been used as standard and effective treatments for CDI for over 25 years although both acute mortality and relapses occur. In horses, metronidazole has been the standard treatment although metronidazole-resistant strains were commonly found in one study [32]. Thus, there is much interest in developing alternative strategies for preventing and treating C. difficile-associated disease. There are numerous reasons for this. First, vancomycin and metronidazole can further disrupt and suppress the return of a healthy intestinal flora, do not reduce exposure to spores in the environment, and do not positively alter host risk factors. Furthermore, C. difficile spores can easily germinate and produce toxins during an approximately 3-month period that is required to reestablish the normal flora of the human intestine after antibiotic treatment [25]. It is also well known that toxin-mediated diseases typically require the production of toxin-specific antibodies for protection. Additionally, antibiotics for CDI target the C. difficile organism but not the toxins that are the actual virulence factors which cause the disease. The use of vancomycin may also promote vancomycin resistance, particularly among enterococci and staphylococci. Furthermore, significant rates of treatment failure and CDI recurrence are associated with metronidazole use in humans. This may be due to increasing metronidazole resistance among C. difficile isolates [30]. Finally, the incidence and mortality associated with CDI are increasing. All of these factors contribute to the pressing need for a novel alternative therapeutic technique.
Recently, immunotherapy has become a very promising strategy especially given the increasing incidence of traditional antibiotic treatment failure and newly emerged hypervirulent C. difficile strains [36]. Previously, researchers developed therapeutic antibodies using birds [20], cows [24,38], and sheep [31]. The ability of these antibodies specific for C. difficile toxins to prevent clinical disease and reducing carriage of C. difficile was tested in hamster and gnotobiotic mouse models. However, there are no reports of the use of vaccines or immunotherapeutic agents against equine CDI [3].To develop anti-C. difficile antiserum for the treatment of horses with C. difficile-associated disease, we prepared antisera of equine origin against both C. difficile toxins A and B as well as the whole bacteria. We then examined the prophylactic effects of passive immunotherapy using these antisera in a conventional mouse model.
Go to : 
Materials and Methods
Bacterial strains and culture conditions
C. difficile isolates 101 and 115 of equine origin were used for this study [34]. Isolate 115 was used to prepare the toxoid because this strain produces a high level of toxin. Isolate 101 was used to challenge the mice. Bacteria were cultivated in pre-reduced anaerobically sterilized peptone yeast extract broth with 1% glucose (Anaerobe Systems, USA) at 37℃ for 48 h under anaerobic conditions.
Animals
4 weeks old female C57BL/6 mice were purchased from Jackson Laboratory (Bar Harbor, USA) for experiments. All mice used in the experiments were housed in groups of five per cage under the same environmental and husbandry conditions. The animals were fed a standard rodent diet (Harlan Laboratories, USA). Food, water, bedding, and cages were all autoclaved. Experimental procedures commenced after 1 week of receipt. All animal experiments were performed with the approval of the Institutional Animal Care and Use Committee (IACUC) at Cornell University.
Preparation of C. difficile spores
Sporulation of C. difficile was induced as previously described [35]. Briefly, an overnight culture of C. difficile was diluted in fresh brain heart infusion (BHI) broth (BD Diagnostics, USA) to an optical density measured at 600 nm (OD600) of 0.2. Next, 150 µL of this suspension was spread on 5 mL BHI agar (BD Diagnostics) in each well of a six-well tissue culture dish (Corning, USA). The culture was incubated anaerobically at 37℃ for 7 days. Each well of the six-well dish containing mixed populations of spores and vegetative cells was flooded with ice-cold sterile water. The vegetative cell-spore mixture was collected, washed five times with ice-cold water, and heated to 60℃ for 30 min to kill vegetative cells. The resulting solution contains the finial spores. 1 mL of the finial spores was diluted from 1 : 103 to 1 : 109 with PBS, plated on BHI agar plates containing 0.1% taurocholate (Sigma, USA), incubated for 2 days and then the resulting colonies were counted. The spores were store at -80℃ until use.
Preparation of C. difficile toxoid
A crude culture filtrate of C. difficile toxoid was used in this study. After C. difficile was cultured inside a dialysis tubing (Thermo Scientific, USA) containing BHI broth for 3 days at 37℃, the supernatant was harvested by centrifuging twice at 10,000 × g for 10 min at 4℃ and concentrated by ultrafiltration through a 50 kDa membrane (Thermo Scientific) using an Amicon concentrator (EMD Millipore, USA). The concentrate was re-suspended in 200ml PBS and concentrated three more times. The last volume of the concentrate was then lyophilized using Freeze Dry System (Labconco, USA) and resuspended in 5 mL PBS. The resulting re-suspended lyophilized culture supernatant was treated with 1% formaldehyde for 10 days at 37℃ and stored at 4℃ until use. Toxin inactivation was identified by the absence of cell culture cytotoxicity relative to treatment with the native toxin as a positive control which caused CACO-2 cells become round while the toxoid didn't.
Bacterin preparation
Bacterin was prepared as previously reported [37]. Briefly, C. difficile was grown in BHI broth at 37℃ for 24 h under anaerobic conditions. The culture was centrifuged at 8,000 × g for 10 min at 4℃ and washed three times with PBS. The pellet was then re-suspended in 500 mL PBS containing 1% formaldehyde and incubated at 40℃ for 24 h. The excess formaldehyde was removed by washing with PBS three times. After the final wash and re-suspension in 100 mL PBS, the number of bacterial cells was determined at OD550 using. The bacterin was cultivated in BHI broth at 37℃ for 24 h under anaerobic conditions to check for live organisms and then stored at -4℃ until use.
Production of antibodies against the bacterin and toxins
Two healthy adult draft horses (1 and 2) were immunized with bacterin and toxoid. Each of the preparations used for immunization were combined with either complete Freund's adjuvant (CFA; Sigma) or incomplete Freund's adjuvant (IFA; Sigma) at the minimum ratio (generally 1 : 1 [v : v]) to create a stable emulsion. The first two rounds of immunization with either bacterin or toxoid were mixed with CFA. All other immunizations with toxoid and bacterin were mixed with IFA. The dose and frequency of each antigen administration is outlined in Table 1. All immunizations were administered via an intradermal (ID) or subcutaneous (SQ) route at multiple sites with no less than 0.1~0.2 mL per site.

SDS-PAGE and Western blotting
To measure the production of antibodies against the toxins, sera collected from the immunized horses were subjected to a Western blot analysis using recombinant toxin A and B fragments prepared in our lab (unpublished) as antigens. These fragments include the C-terminal of TcdA (residues 1820-2710, 127 kDa), the N-terminal of TcdA (residues 1-566, 89 kDa), the C-terminal of TcdB (residues 1820-2366, 89 kDa), and the N-terminal of TcdB (residues 1-566, 90 kDa). The methods used for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting have been previously described [40]. Briefly, 100 ng the recombinant toxin fragments were boiled for 5 min in SDS sample buffer (0.15 M Tris [pH 6.8], 1.2% SDS, 0.3% glycerol, 0.15% β-mercaptoethanol, 0.018% [w/v] bromophenol blue; Sigma) and loaded onto a 12% SDS-PAGE gel to separate the proteins. After electrophoresis, sera collected from the horses were used to detect the proteins transferred to nitrocellulose membranes (Schleicher and Schuell Biosciences, USA) using a semi-dry method [40]. Basically, the membrane was blocked using 5% blotting grade non-fat dry milk (Bio-Rad, USA) in 0.1 M 1× PBS for 60 min at 37℃. The proteins were then detected by incubation with antiserum collected from an immunized animal at a 1 : 200 dilution in blocking buffer at 37℃ for 60 min. The membrane was then incubated at 37℃ for 60 min with 2 µL mouse anti-horse IgG diluted 1 : 3, 000 in blocking buffer as the secondary antibody. Finally, proteins were visualized by Novex AP Chromogenic Substrate (BCIP/NBT; Life Technologies, NY) according to the manufacturer's instructions.
Enzyme-linked immunosorbent assay (ELISA)
An ELISA was used to measure the levels of IgG against the toxins and bacterin. Blood samples were collected before (pre-immune) immunization as well as 11 and 17 weeks after the first immunization. Purified TcdA (List Biological Laboratories, USA), the C-terminal end of TcdB, or a suspension of formalin-killed C. difficile were used to coat 96-well microtiter plates (100 µL/well; Corning) at a concentration of 1 g/well in Bicarbonate/carbonate coating buffer (100 mM). The plates were then blocked with 1% bovine serum albumin (Sigma) in PBS at 37℃ for 2 h. The pre-immune and immune sera were diluted at 1 : 500 in PBS containing 1% bovine serum albumin and 0.05% Tween 20, added to the wells at 100ul/well, and incubated for 1 h at 37℃. IgG reactivity was detected with 1 : 2,500 diluted peroxidase-labeled anti-horse IgG (KPL, USA) for 1 h at 37℃ and then 100 µL of freshly prepared tetramethylbenzidine (TMB) 2-component microwell peroxidase substrate (KPL) were added to each well. Finally, 100 µL of TMB stop solution (KPL) was added to the wells and absorbance of the plates was read at OD450 with a microtiter plate reader (BioTek Instruments, USA).
Cytotoxicity inhibition assay
To assess in vitro neutralizing activities of the prepared horse serum, a cytotoxicity inhibition assay was performed as previously reported [10]. Briefly, 200 ng of purified C. difficile TcdA in modified Eagle's minimum essential medium (Sigma) containing 2 mM L-glutamine, 0.1 mM nonessential amino acids, 1.5 g/L sodium bicarbonate, and 1.0 mM sodium pyruvate was mixed with an equal volume of prepared immune serum or pre-immune serum and incubated for 1 h at 37℃. Caco-2 cells (American Type Culture Collection, USA), which were grown in monolayers, were then incubated with 5 µL mixture at 37℃ for 18 h, and the cytopathic effects were analyzed under a phase-contrast microscope (Nikon, Japan). Untreated cells were used as the negative control and cells treated with 1ug purified C. difficile TcdA prepared by our lab (unpublished) were used as the positive control.
Mouse model of CDI
A mouse model of CDI was established as previously described [8]. Briefly, C57BL/6 mice were given with water mixed with a mixture of kanamycin (0.4 × 10-3 mg/L; Sigma), gentamicin (0.035 × 10-3 mg/L; Sigma), colistin (850 U/mL; Sigma), metronidazole (0.215 × 10-3 mg/L; Sigma), and vancomycin (0.045 × 10-3 mg/L; Sigma) for 3 days. All mice were then given regular autoclaved water for 2 days and received a single dose of clindamycin (10 mg/kg; Sigma) intraperitoneally 1 day before C. difficile challenge. After treatment, all mice were challenged with 107 of prepared toxigenic C. difficile spores by gavage feeding. The animals were observed daily after challenge for the presence and severity of diarrhea and other general symptoms of illness or mortality. Body weight was measured once a day at the same time each day. Mice judged to be in a moribund state were euthanized by CO2. Tissue samples including intestine, live and kidney were collected for histopathologic analysis after euthanasia.
To confirm C. difficile as the causative agent of disease, perianal swabs were taken randomly 2 days after challenge from representative symptomatic mice and cultured anaerobically at 37℃ for 2 days on BHI agar plates containing 5 × 105 mg/L clindamycin. Stool specimens were also collected 2 days after challenge and tested using a C. difficile toxin A/B microplate assay (Remel; Thermo Fisher Scientific, USA) to confirm the presence of toxins A and B. In addition, tissue samples including intestine, live and kidney were taken from representative symptomatic mice, fixed in 10% neutral buffered formalin, sectioned (4-µm-thick) by tissue section machine (Leica, German), stained with hematoxylin and eosin (Sigma), and examined by light microscopy (Nikon, Japan) to confirm the presence of typical CDI-associated lesions.
Treatment of the mice with immune serum
Normal pre-immune or immune serum from the horses was administered intravenously into the tail vein of different groups of mice 1 day before C. difficile challenge. For experiment 1, the protective properties of antisera prepared from the two different horses (1 and 2) named serum 1 and serum 2, respectively, were tested in the mouse model. Mice receiving the immune serum were divided into two groups of five mice each. One group of mice was treated with 100ul immune serum prepared from horse 1 while another was given 100ul immune serum prepared from horse 2. One group of five mice was given normal pre-immune serum (100 µL) from horse 1 alone as a negative control.
For experiment 2, the efficacy of serum from horse 1 was evaluated at two different doses. Two groups of five mice received either 50 µL or 100 µL of the hyperimmune serum prepared from horse 1. Another group of five mice was treated with normal pre-immune serum (100 µL) from horse 1 alone as a negative control. Additionally, one more group of five mice was treated with antibiotics and not challenged to determine whether the antibiotic treatment caused diarrhea or weight loss. Experiment 2 was repeated in duplicate. All the other mice were challenged with C. difficile spores as described above (see "Mouse model of CDI").
Statistical analysis
Differences in mean relative weight loss between the experimental and control mice were evaluated with a two-tailed Student's t test using GraphPad Prism (GraphPad Software, USA). A p value < 0.05 was considered statistically significant.
Go to :
Results
The immunized horses produce antibodies against both toxins A and B
No adverse effects associated with the immunization were observed in the horses. Western blot analysis of postimmunization serum samples from horse 1 showed that the antiserum produced by the immunized horse recognized all recombinant toxin A and B fragments (Fig. 1).
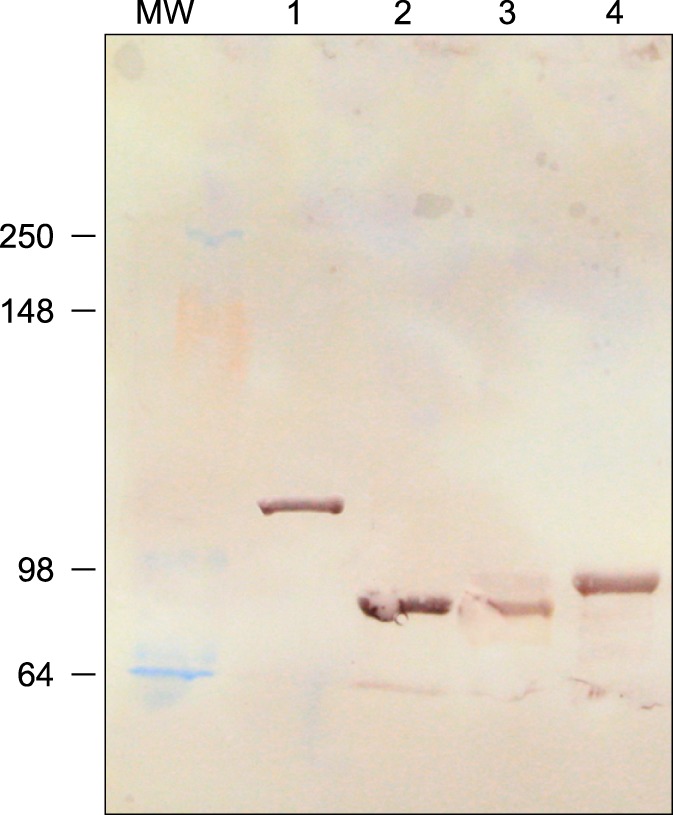 | Fig. 1Equine hyperimmune serum detects purified recombinant N- and C-termini of toxins A and B by Western blot. Proteins were separated on a 12% polyacrylamide gel and transferred to a nitrocellulose membrane (130 V for 90 min.) for the test. The test serum reacted with the purified recombinant N- and C-termini of toxins A and B. Lane MW, molecular weight marker; Lane 1, C-terminal of TcdA (127 kDa); Lane 2, N-terminal of TcdA (89 kDa); Lane 3, C-terminal of TcdB (89 kDa); Lane 4, N-terminal of TcdB (90 kDa).
|
The levels of antibodies generated against C. difficile toxins increased over time
Immune serum samples were collected 11 and 17 weeks after the first immunization from each horse and evaluated with an ELISA. The results showed that the OD values for different sera against purified toxins A and B as well as whole-cell C. difficile continually increased. At the end of immunization protocol, the OD values ranged from 1043 to 1246 for TcdA, 1313 to 1417 for TcdB, and 1730 to 1815 for the whole-cell bacterin. However, the OD values for pre-immune serum were only 65 to 176 for TcdA, 167 to 246 for TcdB, and 534 to 596 for C. difficile bacterin (Figs. 2A-C).
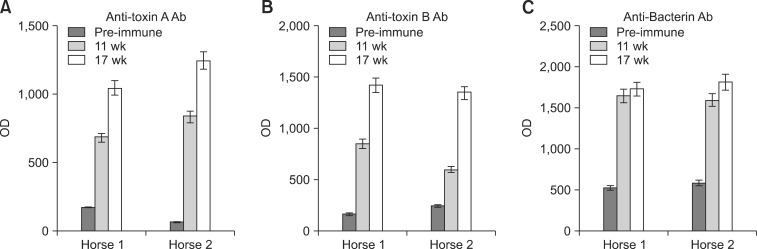 | Fig. 2Antibody titers against TcdA, TcdB, and whole-cell C. difficile bacterin measured by an ELISA. Horses were immunized with C. difficile toxoid or bacterin by sequential intradermal and subcutaneous injection with complete Freund's adjuvant or incomplete Freund's adjuvant. Sera were diluted 1 : 500 and the results are expressed as optical density (OD) units (mean ± SD of duplicate assays). (A) Anti-C. difficile TcdA activity of immune sera measured by an ELISA. (B) Anti-C. difficile TcdB activity of immune sera measured by an ELISA. (C) Anti-C. difficile bacterin activity of immune sera measured by an ELISA.
|
The hyperimmune serum has toxin-neutralizing effects
The ability of the prepared serum to neutralize C. difficile toxin was assessed with a cytotoxicity assay. Cytotoxicity was observed by rounding of Caco-2 cells grown in monolayers after exposure to C. difficile toxins. Controls for this assay included cells treated with the pre-immune serum that lacked specific antibodies against C. difficile toxin A or B as determined by an ELISA. Results of the experiment showed that cells exposed to TcdA after being pretreated with normal pre-immune horse serum became rounded after 24 h of incubation (Fig. 3A). In contrast, pretreatment with antitoxin serum prior to TcdA exposure inhibited cytotoxicity (Fig. 3B). The cells treated with purified TcdA only displayed typical signs of cytotoxicity while the untreated cells showed no signs of toxicity (data not shown).
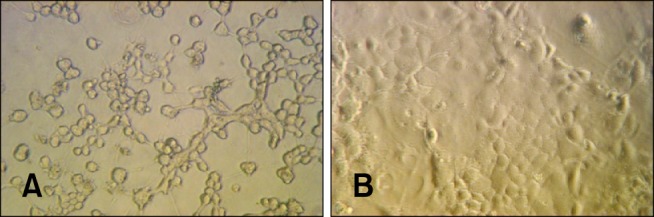 | Fig. 3Toxin-neutralizing activity of the prepared antitoxin serum. (A) Photomicrograph of a Caco-2 cell monolayer pretreated with pre-immune serum for 18 h and exposed to C. difficile toxins. A marked cytopathic effect appearing as rounding of all cells was observed. (B) A Caco-2 cell monolayer pre-incubated with antitoxin serum prepared from hyperimmunized horses and exposed to C. difficile toxins. The monolayer had a normal morphology.
|
Intravenous administration of hyperimmune serum protects mice from C. difficile infection
In experiment 1, the protective effects of antisera prepared from sera of two different horses (serum 1 and 2) were tested in the mouse model. Both serum 1 and 2 given by intravenous injection 1 day before C. difficile challenge protected the mice against severe clinical disease. The group that only received pre-immune serum lost more than 12% body weight on day 3 post-challenge and all animals in this group were visibly sick with diarrhea. In contrast, the mice that received prophylactic antitoxin serum only lost 2% body weight without any signs of illness (Fig. 4A). On day 3, the pre-immune serum control group had lost a significantly greater amount of weight compared to mice treated with serum 1 or 2 (p < 0.05).
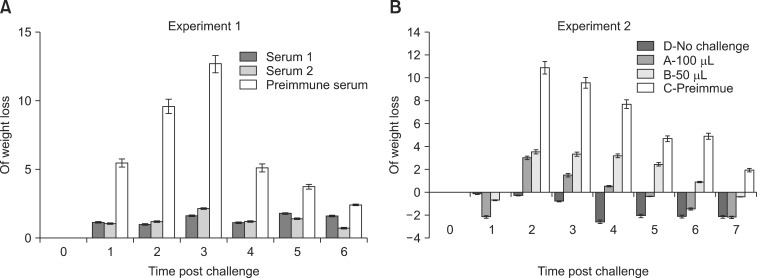 | Fig. 4Relative weight loss of mice challenged with C. difficile after intravenous delivery of hyperimmune serum. The control group had significantly greater weight loss compared to the antiserum-treated groups (p < 0.05). Weight loss of the control mice reached a maximum (> 10%) on days 2 or 3 and returned to normal after day 7. In contrast, weight loss of the antiserum-treated mice was significantly less after C. difficile inoculation and returned to a normal range more quickly compared to the control group. The animals not challenged with bacteria maintained a consistent weight within the normal range. Data are expressed as the mean relative weight loss and standard error for five mice. (A) Mice treated with antitoxin serum from different horses (serum 1 and 2). (B) Mice treated with different doses of antitoxin serum 1.
|
After the preliminary test in experiment 1, the efficacy of serum 1 delivered at two different doses was evaluated in experiment 2. All animals challenged with C. difficile lost weight. The loss of body weight reached a maximum at day 2 and gradually recovered thereafter in all mice. By day 2, animals in the groups that received normal pre-immune serum had lost the most weight (> 10%). The animals that received antitoxin serum lost significantly less weight (p < 0.05) compared to the group that received pre-immune serum alone. Furthermore, the level of protection was dose-dependent. Mice receiving 100 µL of antiserum lost a minimum amount of weight (3.0%) compared to the animals treated with pre-immune serum (p < 0.01) after C. difficile challenge without developing diarrhea. The body weight of these mice also returned to a normal range first (between days 4 and 5 after infection). The group that received 50 µL of antiserum lost more weight (3.5%) compared to mice treated with 100 µL of antiserum. However, this weight loss was still significantly less (p < 0.01) compared to the mice treated with pre-immune serum. Furthermore, it took 2 more days for the body weight of animals treated with 50 µL of the antiserum to return to the normal range than the 100 µL-treated group. By day 7 after infection, the weight of all animals intravenously administered the hyperimmune serum returned to a normal range while the group that received pre-immune serum alone still had a 1.9% weight loss. However, diarrhea in all animals belonging to both the control and antiserum-treated groups had resolved at the time of euthanasia (7 days post-infection). No typical gross or microscopic lesions associated with C. difficile enteritis were observed at the end of experiment (data not shown). The control group that was not challenged with C. difficile consistently maintained their body weight within normal ranges and did not show any sign of disease (Fig. 4B).
Infection was induced by C. difficile
Cultures of perianal swabs from representative symptomatic mice produced large, slightly flat, circular colonies with a filamentous edge. Under the microscope, long, thin, straight Gram-positive rods with typical C. difficile morphology were observed. Additionally, the fecal specimens were positive in the C. difficile toxin A/B microplate assay that confirmed the presence of toxins A and B (data not shown). Histopathological results (Fig. 5) for representative symptomatic mice from experiment 1 showed that these animals suffered from C. difficile-mediated enterocyte apoptosis and/or necrosis with villous atrophy and inflammation based on characteristic changes in the intestine. The presence of microvesicular hepatic lipidosis indicated either acute liver injury or rapid mobilization of peripheral fat stores. Changes in the spleen likely reflected breakdown of the intestinal mucosal barrier and delivery of antigens to the white pulp with early secondary follicle formation. The apparent depletion of zymogen granules in the pancreas may have been reflecting atrophy secondary to anorexia.
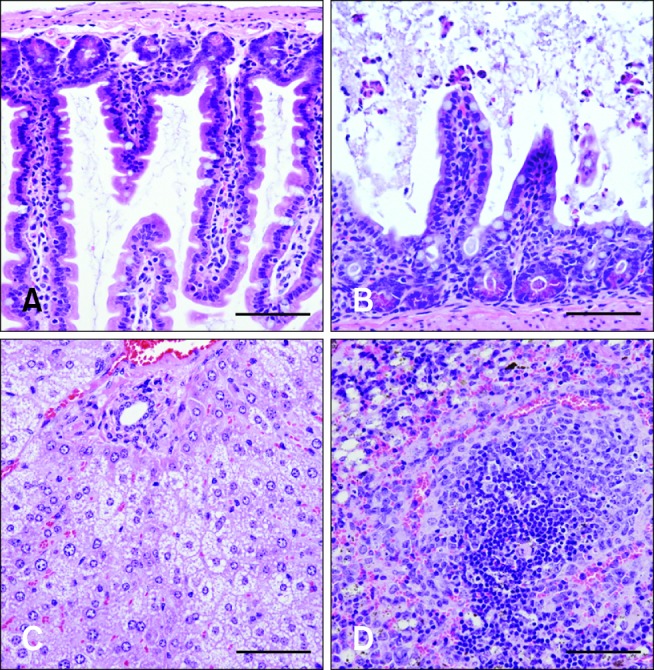 | Fig. 5Histopathology of mice with CDI. (A) Segment of the small intestine from the control mice. (B) Affected segment of the small intestine. Necrotic enterocytes were sloughing from the villous tips and the villi were about 25% shorter than those in the unaffected segment. (C) Microvesicular lipidosis was present in hepatocytes throughout the liver. (D) An early germinal center composed of centroblasts was adjacent to the periarteriolar lymphoid sheath around a nodular arteriole. Note the numerous apoptotic bodies. Hemosiderin-laden macrophages were present in the red pulp. Scale bars = 100 µm.
|
Go to :
Discussion
CDI is a leading cause of nosocomial infections and human health care-associated diarrhea worldwide [36]. In horses, C. difficile is an important cause of diarrhea that leads to sporadic and epidemic disease of varying severity [2]. In a hospital study, horses with diarrhea and that were positive for TcdA had more severe clinical disease than horses with diarrhea without evidence of C. difficile infection [32]. The standard treatment of CDI has focused on the use of antimicrobials such as vancomycin and metronidazole in accordance with antibiotic stewardship guidelines [25]. Although these treatments currently remain the most effective method for controlling C. difficile, immunotherapy techniques undergoing clinical trials involving animals and humans have produced promising results [7,18,39]. Immunotherapy, which involves the administration of antibodies against C. difficile toxins and the transfer of immunity with intravenous immunoglobulins (IVIG), is intended to increase the relatively low level of serum antibodies specific for C. difficile toxins in patients affected by C. difficile. Similarly, immunoprophylaxis could be used to prevent disease in patients at high risk of developing C. difficile-associated diarrhea.
High-risk horses are believed to be those receiving antibiotics known to be associated with C. difficile-related diarrhea and housed in an environment that has had documented problems with this condition [32], or mares that are in contact with macrolide-treated foals [6]. So far, the use of either vaccines or immunotherapeutic sera to prevent or treat this important cause of colitis in horses has not been reported. Over the past decades, equine serum products have become important effective therapeutic agents that are accepted by most equine practitioners and widely used in equine clinics. Polyclonal antibodies are especially effective for neutralizing and clearing toxins from the bloodstream; this is remarkable as there is a significant level of sequence diversity among members of large toxin families [31]. Recently, Artiushin et al. [3] found that immunization of pregnant mares with receptor-binding domains of toxins A and B can lead to the generation of specific antibodies in serum and colostrum that can block the cytopathic activity of the toxins in vitro. With the goal of providing an effective strategy for developing a prophylaxis for CDI in horses, we produced antibodies of equine origin with toxin-neutralizing activity and demonstrated their efficacy in vivo to prevent CDI induced by C. difficile of equine origin.
The ability to produce antitoxin antibodies may be a key determinant for C. difficile colonization versus active CDI. Kyne et al. [22] found that asymptomatic carriage of C. difficile is strongly associated with an immune response to C. difficile toxins that manifests as high serum levels of IgG against TcdA. Patients colonized by C. difficile with low levels of serum IgG against TcdA had a much greater risk for C. difficile-related diarrhea than patients with higher levels of anti-C. difficile IgG [22]. Two other studies also observed the same relationship between high antitoxin A antibody levels and asymptomatic CDI patients [17,26]. More recently, Leav et al. [23] demonstrated that serum antitoxin B antibody correlates with protection against recurrent CDI. Patients with adequate levels of IgG may be more likely to remain asymptomatic carriers. In addition to the protective effect of antitoxin antibodies, anti-surface layer protein (SLP) antibodies against C. difficile have also been found to modulate the course of CDI and enhance the phagocytosis of C. difficile [27]. Therefore, we used both C. difficile toxins A and B along with whole-cell bacteria as antigens when preparing antiserum of equine origin in the current study to provide the greatest potential protection.
Serum polyclonal antibody therapy has been clinically effective for treating many cases of human diseases, but there are still some adverse effects that are commonly associated with IGIV therapy using concentrated immune globulins. Most adverse effects including headaches, flushing, fever, chills, fatigue, nausea, diarrhea, blood pressure changes, and tachycardia are mild and transient. Unfortunately, there is very little knowledge about the mechanisms underlying adverse reactions to IVIG. A small amount of aggregated IgG might be responsible in some patients. IVIG can also interact with microbial antigens in the recipient, leading to the formation of immune complex. In addition, plasma-derived vasoactive or hemodynamically active contaminants can cause some adverse IVIG effects, making it difficult to make dose recommendations based upon safety. Our data showed that a higher dose of serum provides more protection to the recipients; however, higher doses may not necessarily be desirable and could cause unexpected complications considering the potential adverse effects associated with IGIV. An appropriate dose of serum is a crucial factor for the successful clinical use of IVIG. In the current investigation, we used normal but hyperimmunized equine serum rather than IgG concentrate. Similar products have been used in horses and rarely produced adverse side effects [12,15,29].
Animal models of CDI are essential to investigate the use of IVIG for treating active cases of CDI or as a prophylaxis. A hamster model has been used for 3 decades to study anti-CDI therapy and the underlying mechanisms of disease. However, this model only represents a fulminant and lethal disease course, and as such does not represent the usual course and spectrum of CDI in other animals [14,19,20,24]. Gnotobiotic mice can also be colonized by C. difficile and develop intestinal pathology, but the fact that no other microorganisms are present in the intestine is a major drawback [9]. Recently, Saito et al. [33] reported that 100% of animals survived if intravenous immunoglobulin was administered at the same time as C. difficile toxin in a conventional mouse model. However, the mice used in that study were not infected by live bacteria but given a preparation of C. difficile toxin instead. The C. difficile toxin preparation was also administrated by systemic injection rather than via the intestine. In addition, no titer data is available for the antibodies against C. difficile toxin A or B in that study, and this inevitably decrease the clinical value of the study findings as noted by the authors [33].
A good animal model can facilitate much-needed advancement in C. difficile research. Recently, Chen et al. [8] established a mouse model of antibiotic-induced C. difficile-associated disease (CDAD). The key components of CDAD are all mirrored in this antibiotic-induced C. difficile-associated colitis murine model including involvement of the entire colon, pseudomembrane formation, variable disease severity, disease resurgence after vancomycin therapy, and the development of resistance to recurrent disease in some animals that survive an initial episode. All these features offer new opportunities to evaluate therapeutic agents in a model that replicates naturally acquired disease and increase the applicability of the model. In the present study, we examined the protective efficacy of the antiserum against CDI infection using this newly established CDI murine model. The results showed that intravenous antitoxin immunoglobulin administration is an effective method for preventing C. difficile infection in these animals.
Development of C. difficile vaccines has been ongoing for over 2 decades with most based on inactivated toxoid or the recombinant C-terminal cell-binding domain. The efficacy of all these vaccines has been demonstrated in animal models as well as human subjects [11,13,16,28,37]. However, seroconversion after vaccination takes time. In humans, all participants (adults and elderly individuals) receiving the 50-µg dose of vaccine seroconverted to TcdA by day 56 and 75% of all participants seroconverted to TcdB by day 70 [11]. In contrast, antisera can provide protection against CDI as late as 24 h before challenge as shown in our current research. This will provide a great benefit to high-risk patients during emergency situations. High-risk individuals may also receive pretreatment to prevent C. difficile infection before they undergo elective procedures. Moreover, our antiserum might also be useful for treating horses with CDI. We will test this in the next phase of our investigation. Among people with CDI, 20% of patients relapse and 65% of those experiencing a second relapse become chronic cases [1]. However, horses do not relapse or develop chronic disease [1]. We did not determine how long the injected antiserum persists in the recipient's circulatory system, but one injection that provides protection for only a limited period of time may be sufficient in the equine clinic.
In summary, our experimental results provide crucial evidence that immunotherapy is a promising strategy for preventing CDI when administered at an appropriate dose to high-risk patients. This study and previously published investigations have also demonstrated the protective benefit of hyperimmune serum. Based on our findings, further studies about the use of hyperimmune serum in horses are warranted.
Go to :
 XML Download
XML Download