

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Paratuberculosis is a chronic infectious disease caused by Mycobacterium avium subsp. paratuberculosis (Map), an intracellular, slow-growing bacterial pathogen. Paratuberculosis mainly affects domestic and wild ruminants, is a generally sub-clinical condition, and characterized by a long incubation period. Clinically affected animals develop granulomatous enteritis, diarrhea, and loss of body weight, eventually leading to death or culling. The disease is present worldwide and considered a production-limiting factor in domestic ruminants [7]. Paratuberculosis decreases milk production as well as the growth and productive lifetime of cattle [14]. Additionally, an association between Map and Crohn's disease in humans has been suggested [13], raising the concern in the livestock industry of potential trade barriers or decreased consumption of animal products [7].
A diagnosis of paratuberculosis is based either on detection of the causative agent or immune response of the host. Tissue or fecal culturing is considered the most accurate diagnostic test available. However, the use of this diagnostic method has been hampered by costs that greatly exceed those of other diagnostic tests such an Enzyme-Linked ImmunoSorbent Assay (ELISA). One way to overcome the costs associated with fecal culturing for individual animals and minimizing the probability of false-positive results (at the herd level) is to pool fecal samples. The lack of a highly reliable diagnostic test for measuring Map infection is one of the most significant shortcomings that hinder paratuberculosis control [16].
Previously acquired evidence indicates that culture methods using liquid media have greater analytical and diagnostic sensitivity than counterpart techniques which utilize solid media [1,6]. In addition, bacterial growth can be detected sooner using liquid culture modalities [5,25]. However, confirmation of the organism is more difficult with liquid culture because the appearance of colonies and mycobactin-dependence are not observable, and the growth of other non-pathogenic mycobacteria needs to be identified. Nevertheless, once bacterial growth is detected in the broth tube, acid-fast staining, sub-culturing on solid media, or PCR are options for confirming the presence of Map in a sample. PCR represents a rapid and specific means of confirming Map in broth culture [3,12,15,23] and eliminates the need to visualize Map colonies.
Isolation and DNA purification are key steps for the majority of protocols in molecular biology [10]. For most mycobacteria species, the simplest way to obtain DNA from a mycobacterial suspension for PCR assays is boiling for 10 to 15 min in distilled water [19,21]. Herthnek et al. [9] reported that incubating a bacterial suspension at 99℃ or room temperature results in an insignificant DNA yield, suggesting the presence of free DNA. Moreover, free DNA is probably present in liquid culture suspensions as indicated by Sweeney et al. [20] who were able to detect Map organisms in liquid cultures by direct transfer of culture medium to PCR tubes. Extraction of genomic DNA from Map is challenging since this microorganism has one of the slowest growth rates among members of the genus Mycobacterium, and possesses a robust and waxy cell wall. These characteristics render Map cells difficult to lyse.
Published protocols for mycobacterial DNA preparation and commercially available extraction kits are available for PCR applications [9,22]. For these procedures, DNA for Map PCR testing is harvested by proteinase K digestion, phenol-chloroform extraction, and column purification using commercial kits. These procedures are deemed necessary for the release of DNA from mycobacterial cells and separating DNA from PCR inhibitors that are potentially contained in the culture media. Major disadvantages of methods for harvesting DNA from broth culture for subsequent real-time PCR confirmation of Map are high cost as well as substantial time and labor demands. Due to the lack of a simple protocol for extracting DNA from Map liquid cultures, the goal of the present study was to develop a simple and efficient Map DNA harvesting method based on mechanical Map cell disruption and ethanol DNA precipitation. This novel technique was compared to two established methods.
Materials and Methods
Herds and animal population
A total of 517 dairy cows in 15 herds were voluntarily enrolled in this study. All herds belonged to small dairy operations (< 100 milking cows) with a herd size of between six and 60 milking cows, and were located in nine different counties of the De Los Rios Region of southern Chile. The study population included herds with and without previous a history of paratuberculosis based on clinical records and/or test results provided by the owners. These animals grazed year-round while consuming little or no concentrate, and produced < 100,000 kg of milk per year.
Sampling and testing
Fecal samples from all milking cows (> 2 years old) were collected between October and December 2010. The samples from five animals were pooled. A total of 104 pools were cultured and confirmed to contain Map by PCR after DNA extraction using three different Map DNA harvesting methods: the technique developed in the current investigation, a commercial kit, and a reference protocol previously published in the literature. Detailed descriptions of all three protocols are presented below. All laboratory work was conducted at the paratuberculosis laboratory of the Biochemistry and Microbiology Department, Faculty of Sciences, Universidad Austral de Chile (Chile). Pooled fecal samples were inoculated into ParaTB MGIT medium tubes (Becton, Dickinson and Company, USA) to be cultured in the BACTEC MGIT system at 37℃ for 49 days (Becton, Dickinson and Company) according to the manufacturer's protocols. Tubes informed as positives by the BACTEC MGIT system were removed for DNA extraction and real-time PCR targeting the IS900 insertion element.
Novel DNA extraction method (Universidad Austral de Chile, UACH)
MGIT tubes were inverted three times in order to mix the contents and briefly vortexed. From the middle of the tube, an aliquot of 200 µL was aseptically removed and transferred to 1.5-mL centrifuge tubes (Eppendorf tubes; Sigma-Aldrich, USA) and centrifuged at 5,000 × g for 5 min. 18℃. The supernatant in each tube was discarded and the opening of the tube was briefly touched to a clean soft paper tissue in order to remove the remaining liquid. The pellet was dispersed by pipetting with a mixture of 500 µL lysis buffer (2 mM EDTA, 400 mM NaCl, 10 mM Tris-HCl [pH 8.0], and 0.6% SDS) and 2 µL proteinase K (10 µg/µL; Sigma-Aldrich). The solution was then was transferred to a bead-beating tube (Biospec Products, USA) containing 200 µL of beads (0.1 mm zirconia/silica beads; Biospec Products) and incubated at 56℃ for 2 h with shaking at 600 g. The tubes were then shaken in a cell disrupter (MiniBeadbeater-8; Biospec Products) at 3,200 g for 60 sec and incubated on ice for 10 min. In order to remove foam and beads from the inner walls, the tubes were centrifuged at 5,000 × g for 30 sec. The samples were briefly vortexed to ensure that any DNA adhering to small solid particles was not lost when the lysate was transferred.
All liquid contents from the bead-beating tube were transferred to 1.5-mL microcentrifuge tubes (Eppendorf tubes; Sigma-Aldrich) and 500 µL of 100% ethanol were added. The tubes were left standing for 2 min at room temperature before being vortexed for 5 seconds and centrifuged at 18,000 × g for 5 min. 18℃. The supernatant was discarded and the pellet was washed once in 200 µL 70% ethanol by resuspension and centrifugation under the same conditions as mentioned above. Next, the pellet was resuspended in 50 µL of sterile distilled water. The tubes were placed in a dry heating block (Eppendorf; Germany) at 100℃ for 5 min. The solution was briefly centrifuged at full speed (16,000 × g for 30 sec) to remove any contaminating material. Finally, a 25-µL aliquot of supernatant was placed into a new Eppendorf tube (Eppendorf tubes; Sigma-Aldrich) to be used as a template for PCR.
Comparative performance of DNA extraction methods
Each positive pooled sample from the BACTEC-MGIT system was subjected to three different DNA extraction methods: i) the UACH method (described above), ii) a commercial QIAamp DNA Stool Mini Kit (Qiagen, Germany) designed for extracting DNA from fecal samples based on bead beating, proteinase K treatment, and column purification [8]; and iii) cethyltrimethylammonium bromide (CTAB) method, a protocol based on cethyltrimethylammonium bromide and chloroform purification involving the formation of a complex containing all components of a cell except for the nucleic acids [22].
Real-time PCR
Multiplex PCR targeting the insertion element IS900 of the Map genome was performed. The total reaction volume was 20 µL containing 5 µL of DNA template, 1× TaqMan Universal MasterMix (Roche Diagnostics, USA), 0.2 µM of each primer (Roche Diagnostics), and 0.1 µM of FAM probe (Roche Diagnostics). The sequences of the two primers designed to amplify a 63-nucleotide fragment of the IS900 target gene were gacgcgatgatcgaggag (F) and gggcatgctcaggatgat (R). The reactions were processed in a LightCycler 2.0 system (Roche Diagnostics) under the following standard conditions: one cycle at 95℃ for 10 min, 45 cycles at 95℃ for 10 sec, 60℃ for 30 sec, and 72℃ for 1 sec; and cooling at 40℃ for 30 sec. Negative and positive controls (Mycobacterium avium subsp. paratuberculosis ATCC 19698) were included. A positive DNA extraction control was also included. The system performed a presence-absence assay for which only the measurements of the last cycle were recorded. Real-time PCR curves of normalized fluorescence for FAM exceeding a threshold value of 0.01 at less than 40 cycles were considered positive (cycle to positive [CP]) as long as the curves had a normal and expected shape.
Statistical analysis
The three purification methods were compared according to concordance of the final PCR test results (positive vs. negative) and observed differences between CP values. First, the proportion of agreement beyond chance was estimated by calculating the kappa (κ) value for the overall test results. Differences between paired CP values of two purification methods determined the similarity between the two techniques. It was hypothesized that the difference would not be significantly different from zero. This speculation was tested using Student's t test for paired samples. Additionally, the CP values for two purification methods were plotted against each other. The regression line was estimated to identify significant differences between the regression lines and lines of equality (slope different from 1). Statistical differences were regarded as significant when p values were less than 0.05. Finally, Bland-Altman plots were created to visually assess the relative difference between CP values of two purification methods against their mean [2].
Cost analysis
The costs for each DNA extraction protocol performed in the study were estimated. On average, an individual sample cost US$12 to be analyzed by the commercial QIAamp kit. The cost to process a sample with the CTAB protocol was US$7 and only US$3 with the UACH method. According to the estimated sensitivity of each protocol, cost adjustment of each DNA extraction technique for both individual and total number of samples was calculated.
Results
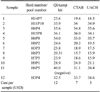
Out of 517 sampled animals (from 15 herds) and 104 fecal pools tested, 12 (12%) pools from eight (53%) herds were positive. The number of sampled animals and pools tested per herd are presented in Table 1. The three DNA extraction-purification protocols produced similar results. Only one significant difference between the three purification methods was observed based on the final PCR results. According to the QIAamp kit, sample 11 (H6P5) was negative while the other two protocols indicated that this sample was positive (Table 2). The cost per processed sample was lowest for the UACH method (Table 2).
Comparison of QIAamp and UACH
The observed mean difference between CP values was 3.8 (95% CI, 1.6~6.0); this difference was significant (p = 0.003). The fitted linear regression equation for the CP values was CPQIAamp = 11.94 + 0.69*CPUACH. The slope (0.69) was significantly less than 1 (p < 0.001), indicating that DNA extraction by UACH was more efficient based on lower CP values. The Bland-Altman plot of differences in CP values between these two purification methods is presented in Fig. 1A. The differences tended to be positive and in favor of the QIAamp kit, although they were fell within the 95% confidence interval (CI) of the agreement region with limits between 10.7 and -3.1.
Comparison of CTAB and UACH
Student's t-test for paired samples identified insignificant differences between CTAB and UACH with a mean difference in CP values of 0.73 (95% CI, -0.48 ~ 1.95). The estimated linear regression equation was CPCTAB ~ 4.29 + 0.86*CPUACH. The slope (0.86) was significantly different from 1 (p < 0.001). As shown in Fig. 1B, the mean difference between CP values was approximately 0, resulting in a narrower 95% CI for the agreement region than the previous comparison (QIAamp and UACH) for which differences between 4.6 and -3.1 were observed.
Comparison of QIAamp and CTAB
Differences between CP values for the QIAamp and CTAB protocols were significantly greater than 0 (p = 0.004) with a mean of 3.06 (95% CI, 1.20~4.92). These data suggest that DNA extraction using CTAB was more efficient than that performed with the QIAamp kit. The regression line was defined by the equation CPQIAamp = 9.27 + 0.77*CPCTAB. The slope coefficient (0.77) was significantly different from 1 (p < 0.001). The mean difference in the Bland-Altman plot (Fig. 1C) deviated from 0, resulting in upper and lower limits of 8.9 and -2.8 for the 95% CI of the region of agreement.
Discussion
In our study, we created a new DNA extraction method (UACH) for confirming the presence of Map with PCR. The goal of our investigation was to develop a simple technique with uncompromised analytical sensitivity and low cost. PCR and statistical analysis results suggested that the UACH method is a useful alternative for harvesting Map DNA from broth culture. This technique is cheaper and less complex than either of QIAamp and CTAB. In the literature, several DNA extraction methods for harvesting DNA from mycobacteria, including phenol extraction protocols, have been described but these procedures commonly involve multiple, time-consuming steps, including the handling of hazardous chemicals [10]. Genomic DNA extraction from Map cells is especially demanding since this microorganism is among the most extreme slow growers belonging to the genus Mycobacterium. Additionally, a robust and waxy cell wall renders Map cells difficult to lyse. Previous studies have evaluated and proposed improvements to methods for Map DNA extraction and purification from liquid cultures for PCR confirmation [12,15,20]. In these investigations, Map DNA was harvested using commercial kits and involved proteinase K digestion along with phenol-chloroform or column purification. These procedures were assumed to be necessary for the release of DNA from mycobacterial cells and separating DNA from PCR inhibitors that may be present in the culture media. In the present study, the DNA extraction protocols we evaluated produced similar results with the exception of sample H6P5 for which the QIAamp method rendered a negative result while those for CTAB and UACH were positive. This false-negative result from the QIAamp kit could most likely be attributed to the elusion column system. This kit was designed to obtain a highly purified DNA template. However, some DNA is lost during the purification process and may potentially affect PCR sensitivity.
It was our intention to assess the effect of the purification protocols on the accuracy of detecting Map infection in the field (southern Chile) rather than for a highly positive or clearly negative laboratory panel. We concede that the number of pools that were positive was relatively low and more robust results would require a larger sample size. However, similar results were produced by one of the standard methods (CTAB) and the one we created (UACH). Additionally, CP values of CTAB and UACH protocols were significantly different from those of the commercial QIAamp kit. This finding could be explained by a lower yield but recovery of more pure DNA when the QIAamp protocol was used. Furthermore, the QIAamp kit is associated with minimal PCR inhibition. Egg yolk is one of the components of the culture media that inhibits PCR [20]. It was removed by centrifuging the samples as proposed by Whittington et al. [24].
Evidence gathered in this study suggests that the UACH protocol represents a better technique than the two reference methods based on similar results and lower cost. This finding coincides with a study in which a protocol for extracting Mycobacterium ulcerans DNA was optimized [11]. For this procedure, a combination of mechanical disruption along with chemical solubilization of the waxy lipid and mycolic acid-containing cell wall was performed to effectively release the DNA. Once the balance between mechanical disruption and chemical solubilization was altered, the DNA quantity and/or quality were compromised.
The key step in the protocol established in the current study is bead-beating that allows a large amount of DNA to be harvested from the bacteria. Purification steps (column-based or chloroform extraction) performed for the QIAamp and CTAB methods are associated with more pure DNA suitable for Map strain-typing methods [4,18]. Bead-beating has been previously performed to lyse Map cells using small zirconia beads [17]. Importantly, pre-treatment with a proteolytic enzyme and lysis buffer helps break tough cell walls with forceful shaking in a cell disrupter (bead-beater). We believe that bead-beating as a means of mechanical disruption along with simple purification and precipitation with ethanol was enough to obtain a higher DNA yield via the UACH method indicated by a lower CP value. The proposed method involved mechanical disruption of the bacterial wall as well as the use of ethanol for purifying and washing the DNA with no need for special or expensive equipment. The technique we have developed will allow diagnostic laboratories to easily extract DNA and improve the ability to accurately detect Map infection.


 XML Download
XML Download