

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
A number of large-scale epidemiological studies have suggested that prolonged and moderate consumption of red wine by the Southern French and other Mediterranean populations is associated with a very low incidence of cardiovascular diseases, despite a high-fat diet, little exercise, and widespread smoking (the so-called 'French paradox') [17]. Although possible compounds responsible for this phenomenon have not been identified, the principal candidates are a number of polyphenolic compounds including stilbenes, flavonoids [6], and resveratrol [19]. Resveratrol; 3,4,5-trihydroxystilbene (RESV) is a natural phytoalexin (phytoestrogen) synthesized in response to injury or fungal attack that is found in grape skin and exists in red wine in its two isomers, trans and cis [19].
Previous studies have shown that resveratrol improves health and extends maximum lifespan by 60% in vertebrate fish [21]. Many studies have demonstrated that resveratrol has a wide range of pharmacological properties, which have been comprehensively reviewed by Bhat et al. [3]. Additionally, resveratrol has been suggested to be cardio-protective via various mechanisms, such as its antioxidant activity [7], inhibition of platelet aggregation [15], induction of nitric oxide (NO) production [10], and modulation of the synthesis of hepatic apolipoprotein and lipids [8].
Recent studies have examined the effects of resveratrol on insulin sensitivity. The results have shown that prolonged intake of resveratrol improves insulin sensitivity and lowers blood glucose levels in high fat diet induced obese mice [2]. Moreover, treatment of high fat diet induced obese mice with high doses of resveratrol reduced body weight and improved mitochondrial functions [12]. Our recent study showed that insulin-stimulated Akt phosphorylation was lower in the liver and white adipose tissue of diet-induced obese mice, but completely normalized by resveratrol treatment [11]. Although resveratrol has various beneficial effects, the detailed mechanisms of resveratrol on the insulin signaling pathway is poorly understood. Therefore, this study was conducted to evaluate the effects of resveratrol on the insulin signaling pathway in a liver of obese mice model. To accomplish this, we administered resveratrol to high fat diet-induced obese mice, then examined the differential levels of protein phosphorylation in their livers using an antibody array.
Materials and Methods
Animal experiments
Animal experiments were performed as described previously [11]. Briefly, male C57BL/6N mice (5 weeks old) were supplied by Orient Co. (Seongnam, Korea). The animals were housed at 24 ± 1℃ and 50% humidity under a 12 h light/12 h dark cycle. All animal experiments were carried out in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals, and were approved by the Institutional Animal Care and Use Committee of Jeju National University (ACUCC; Approval No. 20090018). The mice were fed high fat diets (HF; n = 12) or a normal diet (control; n = 12) for 20 weeks. Additionally, a high fat diet and resveratrol (HFR) fed group (n = 13) was administered resveratrol (30 mg/kg per day by oral gavage) with high fat diet for the last 2 weeks after feeding only high fat diets for 18 weeks.
Measurement of body weights and blood glucose levels
In all animal experiments, body weights were measured once a week. At the end of the experimental period, animals were fasted for 16 h, blood samples were collected from the tail vein, and the blood glucose levels were measured using One Touch Horizon (LifeScan, USA).
Insulin treatment and homogenization
At the end of the animal experiment, mice were fasted for 16 h and then injected with human regular insulin (Eli Lilly and Company, Korea) intraperitoneally at a dose of 5 U/kg to investigate insulin signaling in the liver. The mice were euthanized by cervical dislocation, after which their livers were isolated. Liver pieces (100 mg each) from each mouse (n = 12 per group) were pooled, placed in 5 mL lysis buffer (20 mM Tris-HCl, pH 7.4, with 5 mM EDTA pH 8.0, 10 mM Na2P4O7, 100 mM NaF, 2 mM Na3VO4, 1% NP-40, 13.2 µg/mL aprotinin, 13.2 µg/mL leupeptin, 1 mM PMSF), and homogenized for 90 sec at 11,000 rpm in a tissue homogenizer. The homogenates were then centrifuged at 1,600 × g for 15 min, after which the supernatants were removed and centrifuged at 18,000 × g for 20 min at 4℃. The supernatants of the second centrifugation were stored at -80℃ until used for the antibody array. The protein concentration was measured by the BCA protein assay method (DC protein assay; BioRad Laboratories, USA).
Antibody array
Antibody array experiments were performed according to the protocol recommended by Full Moon BioSystems (USA). Briefly, the antibodies were covalently immobilized on a glass surface coated with polymeric 3-D materials that contain mainly epoxy, aldehyde, and hydroxyl functional groups, allowing covalent attachment of antibodies (Full Moon BioSystems). Each slide consists of an array of antibodies with six replicates per antibody, and multiple positive and negative controls to maximize data quality and reproducibility. A FluoReporter Mini-Biotin-XX-Protein Labeling Kit (Invitrogen, USA) was used for the biotinylation of liver lysate proteins, which was performed according to the manufacturer's protocol. The procedure used to process the array was as follows. The slide was soaked in a Full Moon BioSystems array blocking solution for 30~45 min at room temperature, rinsed extensively with Milli-Q water for 3~5 min, and then dried under a stream of compressed nitrogen. Approximately 70 µg of biotinylated liver lysate diluted in 6 mL of coupling solution from Full Moon BioSystems was applied to each array, after which the samples were incubated for 3 h at room temperature in a chamber with 100% relative humidity. The slide was then washed four to five times with PBS, rinsed extensively with Milli-Q water, and incubated with Cy3-labeled streptavidin solution (0.5 mg/mL) for 45 min at room temperature. This was followed by PBS washing and rinsing with Milli-Q water. After drying by centrifugation, the slide was scanned on a Revolution 4200 microarray scanner (Vidar Systems Corporation, USA), and the images were analyzed with ArraySifter Express 1.3 (Vidar Systems Corporation). The fluorescence intensity of each array spot was quantified, and the mean value and standard deviation of replicates were calculated. The final fluorescence signal (I) was obtained from the fluorescence intensity of each antibody spot after subtraction of the blank signal (spot in the absence of antibody). For each treatment group, a phosphorylation signal ratio induction (Δ) was calculated based on the following equation: Δ=Ip/Inp/I0p/I0np; where Ip and Inp are fluorescence signals of the phosphorylated and non-phosphorylated/total protein from the treated sample, respectively, and the parameters I0p and I0np are those of the untreated control sample. Hence, the phosphorylation signal ratio induction Δ for the control group is given a value of 1. Signals from three different array chips were normalized based on the GAPDH signal after background subtraction.
Results
At the end of the animal experiment, both the HF and HFR groups showed a significant increase (p < 0.01) in their final body weights relative to the control group (Table 1). There was no difference in final body weight between the HF and HFR groups, which suggests that resveratrol does not affect body weight in high fat diet mice. Blood glucose in the fasting state was significantly higher (p < 0.01) in the HF group than the control group, whereas the level decreased significantly (p < 0.05) in the HFR group, indicating that resveratrol reduces blood glucose in high fat diet mice (Table 1).
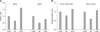
To observe the effects of resveratrol on the insulin signaling pathway in the liver, the phosphorylation levels of proteins on the insulin signaling pathway were observed using an antibody array. At the end of the animal experiment, mice were injected with insulin and livers were isolated from the control group, HF group, and HFR group. Proteins were extracted from the livers of mice of each group, and the phosphorylation levels were analyzed using antibody array. Signals from three different array chips were normalized based on the GAPDH signal after background subtraction (Fig. 1).
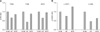
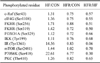
The antibody array revealed that the phosphorylation levels of 10 proteins were increased in the HF group when compared with the levels of the proteins in the control group (Table 2). The phosphorylation levels of c-Raf (Ser43), eIF4G (Ser1108), FKHR (Ser256), FKHR (Ser319), FOXO1A (Ser329), IKKβ (Tyr199), IR (Tyr1361), mTOR (Ser2481), p70S6K (Ser418), and PKCζ (Thr410) in the HF group were increased relative to the levels of the proteins in the control group, whereas the levels were decreased in the HFR group when compared with the levels of the control group (Table 2). The ratio of phosphorylation levels of HFR/HF was lower than 0.7 (Table 2). These results suggest that resveratrol restored the phosphorylation levels of proteins, which were increased in high fat diet mice.
In contrast, the phosphorylation levels of more than 20 proteins were decreased in the HF group when compared with those of the control group (Table 3). The phosphorylation ratios of HFR/HF of the proteins were higher than 1.3 (Table 3). Specifically, the phosphorylation levels of IRS-1 (Ser636/Ser639), PI3-kinase p85-subunit α/γ(Tyr467/Tyr199), and PDK1 (Ser241), which are involved in the insulin signaling pathway, decreased in the HF group, whereas levels were restored to normal levels in the HFR group (Fig. 2). In addition, the phosphorylation levels of Akt (The308, Tyr326, Ser473), GSK-3α(S21) and GSK-3 (Ser9) were restored to normal by resveratrol when compared with the levels in the HF group (Fig. 3). Overall, the results showed that resveratrol restores the phosphorylation levels of proteins involved in the insulin signaling pathway, which were decreased by a high fat diet (Fig. 4).
Discussion
The present study investigated the effects of resveratrol on the insulin signaling pathway in the livers of an obese mice model. Based on the antibody array, the phosphorylation levels of 20 proteins were increased in the HFR group when compared with those of proteins in the HF group. Particularly, the phosphorylation levels of Akt (The308, Tyr326, Ser473) were restored to normal by resveratrol when compared to levels in the HF group. Akt is the major effector of the IR-IRS-1-PI3K pathway, and is activated by phosphorylation of its Thr308 and Ser473 residues; however, the significance of phosphorylation of Akt on tyrosine residues is less well understood. Recent studies have shown that the phosphorylation of Akt on Tyr326 and Tyr474 is required for full activation of the kinase [5,9]. In addition, autophosphorylation of Akt on Ser246 that lie in the characteristic Akt substrate motif (RXRXX(S/T)) appeared to require prior phosphorylation of Akt on Thr308 and Ser473 [13]. The present study showed that resveratrol treatment increased the phosphorylation level of Akt, which is essential for its basal and full activation.
In addition, the phosphorylation levels of IRS-1 (Ser636/Ser639), PI-3K p85-subunit α/γ(Tyr467/Tyr199), PDK1 (Ser241), GSK-3α(S21) and GSK-3 (Ser9), which are involved in the insulin signaling pathway, were decreased in the HF group, whereas levels were restored to normal in the HFR group. One of the approaches used to assess insulin sensitivity involves measuring the ability of insulin to phosphorylate intracellular substrates, such as IRS-1, IRS-2, or Akt, since defects in the activation of these molecules are known to lead to disruption of the insulin signaling pathway [1,16,20]. Recent studies have shown that the phosphorylation of IRS-1 at Ser-636/639 is increased in insulin resistant subjects with impaired Akt activity [22]. PDK-1 is phosphorylated at its activation loop Ser241 residue by autophosphorylation, rather than by an upstream kinase [4]. Once activated, Akt phosphorylates GSK-3α (Ser21) and GSK-3β (Ser9) [14]. The results of the present study show that resveratrol restored the phosphorylation levels of proteins involved in the insulin signaling pathway, which were decreased by a high fat diet.
In contrast, the phosphorylation levels of 10 proteins were decreased in the HFR group relative to the HF group. Specifically, ratios of the phosphorylation levels of HFR/HF of mTOR (Ser2481) and p70S6K (Ser418) were 0.70 and 0.30, respectively. A previous study reported that rapamycin treatment decreased the phosphorylation levels of mTOR at Ser2481 by 30% and almost completely abolished p70S6K phosphorylation [18]. The present study showed that resveratrol treatment decreased phosphorylation levels of both mTOR (Ser2481) and p70S6K (Ser418), suggesting that resveratrol inhibits the mTOR pathway.
Overall, the results showed that resveratrol restores the phosphorylation levels of IRS-1, PI-3K, PDK-1, Akt, and GSK-3, which are the proteins involved in the insulin signaling pathway that were decreased by a high fat diet (Fig. 4). In addition, resveratrol treatment decreased phosphorylation levels of both mTOR (Ser2481) and p70S6K (Ser418), suggesting that resveratrol inhibits the mTOR pathway (Fig. 4). Future studies will be focused on the antibody array of skeletal muscle and adipose tissue isolated from obese mice. Comparison of protein phosphorylation levels among tissues will help elucidate the mechanism underlying the insulin signaling pathway, and will suggest therapeutic approaches for the treatment of diabetes mellitus.




 XML Download
XML Download