

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Alcohol abuse and its associated social consequences are a major health problem in many areas of the world. Alcohol abuse is a brain-centered addictive behavioral disorder that develops regardless of gender, race, age, or economic standing, and can lead to alcoholic liver disease (ALD) in many patients [5,13]. ALD presents a broad spectrum of disorders ranging from simple fatty liver to more severe forms of liver injury that include alcoholic hepatitis (AH), cirrhosis, and superimposed hepatocellular carcinoma [12]. In the United States, the Centers for Disease Control and Prevention estimate that 50% of people aged 18 or older drink alcohol [3]. Among these, 5% are classified as heavy drinkers and 15% are binge drink. The National Institute on Alcohol Abuse and Alcoholism (USA) has reported that liver cirrhosis is the 12th leading cause of death with a total of 29,925 deaths in the United States in 2007 (48% of which were alcohol-related) and is also a major contributor to liver disease-related mortality in other countries [12].
The abuse of alcohol significantly increases morbidity and mortality from infectious diseases along with the risk of cardiovascular, brain, pancreatic, renal, cerebral, and oncological diseases [13]. The overall socioeconomic cost of alcohol abuse including direct (drug and hospitalization expenses) and indirect (due to loss of work productivity, crime, and reduction in health-related quality of life) costs has been estimated to be more than $185 million annually in the United States [20,25]. Despite the profound economic and health impact of ALD, many patients continue to consume alcohol. The primary treatment recommendation for this disease is abstinence from alcohol. Other treatment options, such as pharmacological and nutritional therapies, are also available [17]. Interest in potential antioxidant therapy for treating AH has increased as a growing body of evidence indicates that oxidative stress is a key mechanism underlying alcohol-mediated hepatotoxicity. Alcohol intoxication leads to increased generation of reactive oxygen species (ROS) while suppressing the antioxidant defense system [28,31]. Antioxidants have the ability to scavenge/deactivate ROS and inhibit the oxidation of various cellular substances. Several antioxidants such as S-adenosylmethionine, N-acetylcysteine, beta-carotene, vitamin C, vitamin E, and selenium have been systematically evaluated [12,18]. It was reported that AH patients treated with vitamin E, selenium, and zinc supplements had an in-hospital mortality rate of 6.5% compared to a mortality rate of 40% for the placebo group [17]. Plants produce an extensive variety of antioxidant compounds such as phenolics, terpenoids, alkaloids, and bioactive protein or peptides. These reagents can be used as ingredients in nutritional supplements/nutraceuticals that combat oxidative stress with minimal side effects [35].
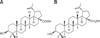
Betulinic acid (BA; 3β-hydroxy-lup-20(29)-en-28-oic acid) is a pentacyclic lupane-type triterpene (Fig. 1A) that exists widely in food, medicinal herbs, and plants, especially birch bark [9]. As an oxidation product of betulin (see Fig. 1B), BA has also been detected in extracts of white birch bark [9,32]. These reagents are of particular interest due to their wide spectrum of biological activities such as antioxidant, anti-inflammatory, anti-tumor, anti-angiogenesis, anti-viral, anti-HIV, anti-neoplastic, and anti-plasmodial properties [10,11,22,24,32]. BA was not found to be toxic at a concentration of 500 mg/kg in mice [22].
Several studies have demonstrated that natural triterpenes such as lupeol, betulin, ursolic acid, and oleanolic acid effectively reduce hepatotoxicity induced by carbon tetrachloride, acetaminophen, ethanol, and cadmium in vivo and in vitro [29,38]. The protective effect of betulin, BA, and oleanolic acid against ethanol-induced cytotoxicity in HepG2 cells has also been reported [28]. The mechanisms underlying this effect include the suppression of enzymes activities that play roles in liver damage such as cytochrome P450 (CYP450), cytochrome B5 (CYB5), and cytochrome P4501A (CYP1A), or increased production of antioxidant substances including catalase (CAT), superoxide dismutase (SOD), glutathione (GSH), and metallothioneins that help protect liver mitochondria [28]. In vitro, BA inhibits ethanol-induced activation of hepatic stellate cells, as a antioxidant, suppressing of ROS, tumor necrosis factor-α (TNF-α) and transforming growth factor-β (TGF-β) production as well as nuclear factor-kappa B/inhibitory protein of NF-κB (NF-κB/IκB) transduction signaling [28,29]. However, ability of BA to confer in vivo protection against liver injury induced by alcohol has not been previously reported. The objective of this study was to investigate the possible hepatoprotective effects of BA against alcohol induced acute liver damage in mice and elucidate the underlying mechanism.
Materials and Methods
Extraction, Synthesis, and Identification of BA from White Birch Bark
White birch bark samples were collected during the spring of 2009 in Wroclaw, Poland. All the collected bark samples were immediately dried at 60℃ and stored in a dry, dark place. To extract betulin, about 15 g of dried bark were subjected to reflux with 200 mL methanol for 3 h at 70℃. The methanol extract was dried under negative gauge pressure and dissolved in 150 mL dichloromethane. After adding 150 mL 2 M sodium hydroxide and mixing, the lower liquid layer was collected and then filtered under negative gauge pressure. The residue was dissolved in 200 mL ether. Next, 200 mL water was added and the solution was thoroughly mixed. The upper liquid layer was subsequently collected, dried, and subjected to silica gel column chromatography (Flash column; Aisimo Corporation, China) [32]. Betulin was fractionated with hexane and ethyl acetate (6 : 1, v : v). BA was synthesized in two steps: 1) the betulin was oxidized with Jones reagent (1 : 6, molar ratio) at 20℃ for 3 h to obtain betulonic acid, and 2) betulonic acid was reduced by sodium borohydride (1 : 1, molar ratio, Sigma-Aldrich, USA) in 10 mL tetrahydrofuran (Sigma-Aldrich) to produce a mixture of 3α- and 3β-hydroxyl products (5 : 95) as previously described [9,32]. Crystallization of the product from methanol resulted in the 3β-hydroxyl product (BA) with a yield of 75%.
The semi-synthetic compound was a white powder, and characterized using Fourier transform infrared spectroscopy (FI-IR), high performance liquid chromatography-mass spectrometery (HPLC-MS), and nuclear magnetic resonance (NMR) as previously described [32]. Purity of the BA was measured with HPLC using a Zorbax Eclipse XDB-C8 column (4.6 × 150 mm, 5 µm; Agilent Technologies, USA). The injection volume was 20 µL and the column was eluted with an ethanol : water solution (86 : 14, v : v, 4 g/L ammonium formate in water) as the mobile phase at a flow rate of 1 mL/min, and UV detection was performed at 210 nm at room temperature. The Purity of the BA was 96.5%.
Animals and Experimental Design
A total of 50 male Kunming mice weighing 18~22 g (6 weeks old) were purchased from Hunan Silaikejingda Laboratory Animal Co., Ltd. (China). The mice were acclimated to the laboratory environment for 1 week before the experiment was initiated, and fed an M02 standard mouse diet from Hunan Silaikejingda Laboratory Animal (China). The mice had access to food and water ad libitum, and were maintained at a constant temperature (23 ± 1℃) and humidity (60 ± 10%) with a 12-h light/dark cycle. The present study complied with the Animal Care and Use Guidelines of China and was approved by the Animal Care Committee of Hunan Agricultural University (China) [15].
The dose of BA and duration of treatment were established based on a previous study [37]. The mice were randomly divided into five groups: group A served as the control (no alcohol, no BA), group B served as the alcoholic liver injury model (alcohol, no BA), group C received a low dose of BA (0.25 mg/kg) and alcohol, group D received a moderate dose of BA (0.5 mg/kg) and alcohol, and group E consumed a high dose of BA (1 mg/kg) and alcohol. BA was administered orally to most mice with 1% starch jelly (Sinopharm Chemical Reagent, China) every day for 14 days while animals in groups A and B were given an equivalent amount of 1% starch jelly only. The mice were fasted overnight (16~18 h) before receiving a single dose of 50% ethanol (Sinopharm Chemical Reagent) at the dosage of 10 mL/kg on the 14th day of the study 1 h after the last administration of BA. Group A received an equal volume of sterile saline.
Nine h after alcohol administration, blood samples were collected in tubes (Eppendorf, Germany) without anticoagulant by venous puncture with the mice under light anesthesia induced by diethyl ether (Sinopharm Chemical Reagent). The animals were then sacrificed by cervical dislocation and liver samples were collected. Serum was collected by centrifugation (Z383K Universal High Speed Centrifuge; Hermle labortechnik, Germany) at 2,000 × g for 10 min at 4℃ and frozen at -70℃ until analysis. Livers were quickly removed, weighed, and the liver indices expressed in milligrams of liver per 10 g of body weight were calculated. The liver tissues were washed in a chilled 0.9% NaCl solution and minced into small pieces on ice. A 10% (w/v) homogenate (Tenbroeck tissue grinders; Wheaton, USA) was prepared in 10 mM phosphate buffer (pH 7.4) and centrifuged (Z383K Universal High Speed Centrifuge; Hermle labortechnik) at 2,500 × g for 15 min at 4℃. The resulting supernatant was collected and stored at -70℃ until analysis.
Evaluation of Serum Biomarkers of Liver Injury
Serum alanine transaminase (ALT), aspartate transaminase (AST), total cholesterols (TC), and triacylglycerides (TG) levels were measured using Mindray commercial kits and a Mindray BS-200 automatic biochemistry analyzer (Shenzhen Mindray Bio-Medical Electronics, China). ALT and AST levels were expressed as units per liter of serum (U/L) while TC and TG concentrations were expressed as nmol per liter of serum (nmol/L).
Measurement of SOD, GSH-Px, and CAT Activities Along with Hepatic MDA and GSH Contents
The activities of superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and catalase (CAT) were measured along with malondialdehyde (MDA) and glutathione (GSH) levels using commercial kits (Nanjing Jiancheng Bioengineering Institute, China) according to the manufacturer's protocols. The levels of SOD, GSH-Px, and CAT activities were expressed as unit per mg of protein (U/mg protein). The MDA and GSH levels were expressed as nmol per mg protein (nmol/mg protein) and mg per g of protein (mg/g protein), respectively. Liver protein concentrations were determined with a Bradford assay using commercial kits (Nanjing Jiancheng Bioengineering Institute) according to the manufacturer's protocols.
Histologic Analysis of Liver Tissues
Liver tissues from the mice were removed and fixed in 10% neutral buffered formalin. The tissues were then rinsed with water, dehydrated with ethanol, subsequently embedded in paraffin, and sectioned into slices (4-µm-thick) using a Leica RM2235 rotary microtome (Leica Microsystems, Germany). The sections were mounted onto glass slides (Beyotime Institute of Biotechnology, China), dewaxed in xylene and ethanol, and stained with hematoxylin and eosin (Beyotime Institute of Biotechnology, China) for histological evaluation using a Motic BA410 microscope (Motic Incorporation, China). Image Pro-Plus Motic Med 6.0 software (Motic Incorporation) was used for image processing.
Statistical Analysis
The results are presented as the mean ± standard deviation (SD). The data were analyzed with a one-way analysis of variance (ANOVA) followed by the Student-Newman-Keuls test for multiple comparisons. All statistical analyses were performed using the SPSS 16.0 statistical package (SPSS, USA). A p value < 0.05 was considered statistically significant and a p value < 0.01 was considered highly significant.
Results
Effects of BA on Liver Indices and Serum Biomarker Expression in Mice
The effects of BA on alcohol-affected liver indices along with serum levels of ALT, AST, TC, and TG in mice were assessed (Table 1). The liver indices of group B were increased (p < 0.05) after alcohol treatment compared to those in the control group A. Pretreatment with BA significantly restored the liver indices of groups C, D, and E in a dose-dependent manner compared to those of group B.
The activities of ALT and AST in alcohol-treated group B were increased by 45.3% and 130.5%, respectively, compared to those found in the control (group A; p < 0.01). Pretreatment with BA reduced both serum ALT and AST levels in a dose-dependent manner. Serum ALT and AST activities were decreased by 7.6% and 9.4% (p < 0.05), 17.8% and 17.0% (p < 0.01), and 25.4% and 30.9% (p < 0.01), respectively, in groups C, D, and E compared to those observed in group B. At the same time, the serum concentrations of TC and TG in group B were increased by 15.7% and 38.7%, respectively, relative to group A (p < 0.05). BA pretreatment reduced the concentrations of TC and TG in a dose-dependent manner. Serum TC and TG levels were decreased by 6.0% and 11.1%, 11.3% and 18.2% (p < 0.05), and 12.8% and 24.5% (p < 0.05), respectively, in groups C, D, and E compared to group B. These findings indicate that BA pretreatment conferred significant protection against alcohol-induced fatty liver by reducing elevated serum levels of ALT, AST, TC and TG in a dose-dependent manner.
Effect of BA on SOD, GSH-Px, and CAT Activities in Mouse Liver
SOD, CAT, and GSH-Px are important enzymes in the antioxidant defense system. Hepatic activities of these three factors decreased significantly in alcohol-treated mice (group B) compared to the control group A (Fig. 2). Pretreatment with BA (groups C, D, and E) increased the activities of SOD, CAT, and GSH-Px in a dose-dependent manner compared to treatment with alcohol alone (group B).
Effects of BA on Hepatic MDA and GSH Levels in Mice
MDA is a major biomarker of lipid peroxidation in tissues. GSH is an antioxidant that helps protect cells against ROS. As shown in Fig. 3, MDA levels was remarkably increased by 87.5% (p < 0.01) while GSH levels were significantly decreased by 46.3% (p < 0.01) in the alcohol-treated animals (group B) 9 h after alcohol administration compared to the control group A. BA pretreatment significantly decreased hepatic MDA contents while increased GSH levels in a dose- dependent manner. MDA contents were decreased by 12.8%, 17.4%, and 39.7% while the GSH levels were increased by 39.4%, 54.8%, and 66.0% in groups C, D, and E, respectively, compared to group B (Fig. 3).
Effects of BA on Mouse Liver Histopathology
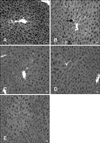
To confirm the protective effect of BA on alcohol-induced liver tissue damage, histological examinations were also performed. The control group had normal hepatic architecture (Fig. 4A) whereas the mice treated with alcohol alone exhibited diffuse cytoplasmic vacuolization (Fig. 4B) indicative of hepatocellular steatosis with no signs of necro-inflammation. In contrast, BA pretreatments reduced the degree of alcohol-induced micro-vacuolization in the liver in a dose-dependent manner (Figs. 4C, D, and E). These results showed that BA protected the mouse livers against alcohol-induced hepatic steatosis, a known indicator of alcohol toxicity.
Discussion
BA is a naturally occurring plant-derived pentacyclic triterpenoid present in many fruits and vegetables such as Tecomella undulata, Coussarea paniculata, Caesalpinia paraguariensis, Vitex negundo, and Ilex macropoda [14,33]. Although BA is widely available from numerous botanical sources, BA contents in these sources are extremely low. Alternatively, BA can be chemically synthesized from betulin, a closely related compound. White bark birch trees (Betula species), a major source of betulin, are widely distributed in the northern hemisphere. Betulin accounts for up to 30% of the dry weight of birch bark extract [9]. Therefore, a semi-synthetic method for preparing BA from betulin was used to obtain sufficient quantities of this bioactive triterpene for the current study.
Alcohol abuse affects the morphology and function of almost all organs in the body [27,31]. Liver is one of the prime target organs of alcohol-related diseases. Alcohol abuse results in hepatocytes damage, liver injury, and inflammation that lead to increased cell permeability and leakage of AST and ALT. Hence, elevated activities of AST and ALT in the plasma are hallmarks of hepatic damage [14,23]. In the present study, alcohol treatment significantly increased AST and ALT activities. These activities significantly decreased when the mice were pretreated with BA. The results were in agreement with previous findings showing that mice pretreated with BA have significantly reduced serum levels of ALT and AST elevated by d-galactosamine/lipoplysaccharide [38], indicating that the hepatocytes sustained minimal injury and their membranes were stabilized.
Alcohol alters the fatty acid composition in the liver. It has been demonstrated that increased cellular nicotinamide adenine dinucleotide hydrogen concentrations and acetaldehyde dehydrogenase activity may impair fatty acid β-oxidation and the tricarboxylic acid cycle during alcohol metabolism, leading to severe free fatty acid overload, increased TG synthesis, and subsequent steatosis [6,27]. Results from the present study showed that BA pretreatment could protect against alcohol-induced fatty liver by reducing serum levels of TC and TG as well as liver indices, which was most likely due to increased β-oxidation in the liver and energy expenditure. BA also ameliorated steatosis (Fig. 4), indicating that BA decreased lipid accumulation in the visceral adipocytes of alcohol-treated mice. This in turn could prevent toxic complications resulting from increased levels of hepatic lipids. Previously, it was also reported that BA functions as both a hypolipidemic and hypoglycemic agent when administered daily to mice fed a high-fat diet [8].
Alcohol affects the liver by inducing nutritional disturbances along with its predominant metabolism in the liver associated with oxidation-reduction (redox) reactions and oxidative stress [26]. In hepatocytes, oxidation of alcohol into water and carbon dioxide is mediated by three known enzyme systems: alcohol dehydrogenase (ADH) in the cytoplasm, the microsomal ethanol oxidizing system in the smooth endoplasmic reticulum of mitochondria, and CAT in peroxisomal membranes [13]. All of these biochemical pathways produce acetaldehyde, a highly reactive product of alcohol oxidative metabolism that damages mitochondria and promotes glutathione depletion, free radical-mediated toxicity, and lipid peroxidation [12]. Alcohol also increases liver oxidative stress via the generation of ROS and adducts. Excessive ROS or a defect of antioxidant system can directly lead to hepatocyte damage and enhance the sensitivity of hepatocytes to lipid peroxidation [27]. Endogenous enzymatic and non-enzymatic antioxidants protect the intracellular milieu against various toxic factors. Elevated lipid peroxidation results in significant depletion of cellular antioxidants [14]. Cellular GSH levels can be decreased through ROS-induced GSH oxidation or GSH export from the cells. Reduced GSH concentrations further enhance ROS production during oxidative challenge [7]. Moreover, BA significantly inhibits ROS production by hepatic stellate cells treated with ethanol [29]. All of these observations indicate that BA exerts prominent antioxidant effects against alcohol-induced oxidative damage.
Oxidative stress results in apoptosis and cell damage, and can directly disrupt the intracellular redox state or indirectly activate signal transduction pathways [37]. Apoptosis is triggered by extrinsic and intrinsic signals via two main pathways: the death receptor and mitochordria-mediated cascades [16]. The death receptor pathway involves ligand-receptor binding initiated by proteinprotein interactions at the cell membranes that activate initiator caspases [1]. Ligands that activate death receptors belong to the tumor necrosis factor (TNF) superfamily of cytokines that includes TNF-α, Fas ligand (FasL), and tumor necrosis factor-alpha-related apoptosis-inducing ligand (TRAIL) [2]. TNF-α, one of the most important proinflammatory mediators, induces hepatocyte apoptosis and necrosis through the activation of caspase-3, thereby leading to hepatocellular DNA fragmentation [36]. Several studies have shown that oxidative stress in cases of alcohol liver injury could result in the up-regulation of TNF-α expression [19,29,38]. TNF-α-induced intracellular ROS generation is markedly decreased by pretreatment with BA [34]. Moreover, BA decreases ethanol-induced TNF-α and TGF-β expression, probably through its ability to inhibit the NF-κB pathway [19,29,38].
B-cell lymphoma/leukemia-2 (Bcl-2) is an anti-apoptotic protein, and Bcl-2-associated X protein (Bax) and Bcl-2 antagonist/killer (Bak) are pro-apoptotic proteins that are important regulators of apoptosis in the mitochondrial pathway [4,30]. BA promotes the expression of Bcl-2 and suppresses acute liver damage by preventing apoptosis [38]. Additionally, ROS activation of c-Jun NH2-terminal kinase (JNK) can induce extrinsic or intrinsic apoptotic signaling. Previous reports have shown that BA significantly inhibits the production of ROS and down-regulates JNK activity in hepatic stellate cells treated with ethanol [21,29]. Therefore, the protective effects of BA against alcohol-induced liver injury possibly involve both exterior and interior pathways.
In conclusion, the present study demonstrated that BA exerts potent hepatoprotective effects that reduce alcohol-induced liver damage in mice. The primary mechanisms responsible for the hepatoprotective activities of BA might be due to the antioxidant capacity of this compound that improve the tissue redox system, maintain the antioxidant system, and decrease lipid peroxidation in the liver. These properties may involve both external and internal pathways. BA has a great potential for use as a beneficial nutraceuticals or dietary supplements for treating alcohol-related diseases. Further investigation is necessary to determine the exact mechanism governing the ability of BA to prevent alcohol-induced liver damage. It is also worth investigating the in vivo and in vitro roles of BA in apoptotic pathways that include signal transduction cascades and pro-apoptotic cytokines.



 XML Download
XML Download