

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Bone marrow-derived mesenchymal stem cells (MSCs) are an excellent model for developing cell-based therapeutics because of their potential for repairing damaged tissue. In particular, MSCs possess advantageous attributes such as ease of isolation, high expansion potential, genetic stability, and multipotent capacity. For these reasons, MSCs have emerged as the most suitable candidate for cellular therapy and tissue engineering techniques. Several preclinical studies have demonstrated that MSCs help prevent deleterious ventricular remodeling and improve the recovery of cardiac performance after myocardial infarction [23,31]. However, Toma et al. [34] reported a very low survival rate (<1%) of MSCs transplanted into ischemic hearts 4 days after transplantation. Poor MSC viability after transplantation critically limits the efficacy of this new therapeutic strategy [23,25,31].
A major challenge to stem cell therapy is that transplanted stem cells undergo apoptosis at an accelerated rate given that they are exposed to an extremely harsh, pro-apoptotic microenvironment in the injured region [10]. In addition to a poor blood supply and low oxygen tension, inflammation triggered by tissue damage may generate oxidative stress as well as cytotoxic radicals and proteins, all of which may cause the death of transplanted cells through several different mechanisms including apoptosis [10]. In particular, hypoxia, defined as an insufficient oxygen supply to tissues, can result from physiological or pathological conditions and changes the physiological characteristics of embryonic and adult stem cells [6]. The mechanism underlying hypoxic condition might be primarily attributed to the hypoxia inducible factor-1 (HIF-1) signal pathway [37]. HIFs control cellular responses to hypoxia by regulating genes that are involved in metabolism, angiogenesis, erythropoiesis, cell proliferation, differentiation, and apoptosis [13]. HIF is a heterodimer consisting of an oxygen-sensitive alpha subunit (HIFα) and a constitutively active beta subunit (HIF-1β; aryl hydrocarbon receptor nuclear translocator). Three isoforms of HIFα have been identified. HIF-1α and HIF-2α are thought to modulate the majority of responses to hypoxia involving HIFα [21]. In the presence of hypoxia, stabilized HIFα binds to HIF-1β, and the resulting complex regulates the expression of downstream genes involved in adaptation to and protection against low oxygen levels [21].
MicroRNA (miRNA) consists of single-stranded noncoding RNA containing 19-23 nucleotides that mechanistically influences a variety of cellular functions via translational inhibition or degradation of target mRNA. Given the diverse roles of miRNA in numerous cellular functions, the role of this molecule in gene regulation associated with hypoxia is not surprising. Recently, miRNA molecules have emerged as a new class of genes regulated by HIF-1α in response to hypoxia. miRNA-199a (miR-199a), miR-17-92 clusters, and miR-20b regulate HIF-1α with hypoxia [19,29,32] whereas the expression of miR-23, miR-24, miR-26, miR-107, miR-210, and miR-373 is induced by HIFs [5,12]. Following the first report of hypoxia affecting miRNA expression by Ivan and colleagues in 2007 [18], a number of studies have demonstrated that miR-210 is the most consistently and predominantly up-regulated miRNA in hypoxic condition. In cancer cells, miR-210 over-expression under normoxic conditions leads to increased HIF-1α levels [13]. Consequently, HIF-1α activity increases, triggering the accumulation of miR-210 as oxygen levels decrease. Increased miR-210 levels suppress the protein expression of glycerol-3-phosphate dehydrogenase 1-like (GPD1L) as a negative regulator of HIF-1α stability and a direct target of miR-210, resulting in increased activity of HIF-1α [14,28].
In the present study, we hypothesized that the positive feedback loop between miR-210 and HIF-1α would improve the survival of human MSCs (hMSCs) under hypoxic conditions. To test this hypothesis, we confirmed the induction of HIF-1α and miR-210 expression in hypoxic hMSCs. We also determined that HIF-1α and miR-210 mutually regulated each other's activities when miR-210 was over-expressed and HIF-1α was down-regulated in MSCs. Moreover, miR-210 overexpression regulated cell survival and modulated HIF-1 activity.
Materials and Methods
Materials
Cobalt (II) chloride hexahydrate (CoCl2), Z-Leu-Leu-Leu-al (MG132), and anti-β-actin antibody were purchased from Sigma-Aldrich (USA). Anti-HIF-1α antibody (H-206) was obtained from Santa Cruz Biotechnology (USA). Anti-phospho-Akt, Akt, anti-phospho-ERK1/2, and ERK1/2 antibodies were obtained from Cell Signaling (USA).
hMSC culturing
hMSCs (cat. No. PT-2501) were obtained from Lonza (USA). The cells had been harvested from normal human bone marrow and were maintained at 37℃ humidified atmosphere containing 5% CO2 incubator. Culture media composed 10% fetal bovine serum (FBS; Invitrogen, USA) Dulbecco's modified Eagle's medium (DMEM)-low glucose supplemented with and 100 U/mL penicillin (Invitrogen, USA) and 100 µg/mL streptomycin (Invitrogen, USA). Fresh media was added and replaced every 3 or 4 days. We used the hMSCs after 7~10 passages. At these passage numbers, hMSCs maintained a consistent morphology and were positive for various mesenchymal markers such as CD29, 44, 105, and 166 by flow cytometry (data not show).
Experimental induction of hypoxia
MSCs were cultured until 80% confluent and were then grown under hypoxic conditions for 12~24 h. The medium used for culturing under hypoxic conditions was serum-free DMEM containing 100 U/mL penicillin and 100 µg/mL streptomycin. The medium was degassed and then exposed to a gas mixture of 5% CO2, 10% H2, and 85% N2. The hypoxic cells were maintained in an airtight humidified hypoxic chamber (Anaerobic Environment; ThermoForma, USA) at 37℃ and continuously gassed with 85% N2. Cobalt chloride (CoCl2; Sigma, USA) was dissolved in PBS and used at a final concentration of 250 µM in serum-free DMEM under normoxic conditions for 24 h.
miRNA transfection
miRNA was purchased from Genolution Pharmaceuticals (Korea). hMSCs were transfected with the miRNA at a final concentration of 100 nM using siLentFect Lipid reagent (Bio-Rad, USA) in serum free DMEM. miR-210 mimic, chemically optimized double-stranded RNA, or scrambled RNA oligomer was transfected into cells. After transfection for 4 h, the media were replaced in 10% FBS contained DMEM.
Real-time PCR
MSCs were lysis using 1 mL TRIzol reagent (Sigma, USA). Total RNA was extracted using chloroform (200 µL) and shaked vigorously for 15 sec and incubate at room temperature for 7 min. Next, centrifugation at 12,000 × g, at 4℃ for 15 min, caused three layers to appear, and the transparent upper aqueous phase was collected in a new tube. Each sample next received 500 µL 2-propanol, and inverted for 10 sec was repeated and centrifugation at 12,000 × g, at 4℃ for 10 min. The pellet washed in diethylpyrocarbonate-contained 75% ethanol. Centrifugation at 7,500 × g, at 4℃ for 5 min, followed. Remove all leftover ethanol and pellet dried about 2 min at room temperature. The isolated RNA was resuspended in 30 µL nuclease-free water. The quantity and quality of the RNA were determined by measuring the OD260/280 ratio with a DU 640 spectrophotometer (Eppendorf, Germany). Next, 100 ng the purified total RNA underwent reverse transcription (Taqman MicroRNA Reverse Transcriptase kit; Applied Biosystems, USA) in combination Taqman MicroRNA assays. The threshold cycle (Ct) of each target gene was automatically defined, located in the linear amplification phase of the PCR, and normalized to control U6 (ΔCt value). The relative difference in expression levels of each miRNAs was calculated (DDCt) and is reported as fold induction (2-ΔΔCt). Amplification and detection of miR-210 or U6 were performed using a TaqMan Small RNA Assay kit (Applied Biosystems, USA) at 95℃ for 10 min followed by 40 cycles of 95℃ for 15 sec and 60℃ for 60 sec in a Light Cycler 480 II (Roche, Switzerland). The threshold cycle (Ct) of each target gene was automatically identified in the linear amplification phase of PCR and normalized to the cycle number of the RNU19 control (ΔCt value). Relative differences in the expression levels of each miRNA sequence were calculated (ΔΔCt) and reported as fold-induction (2-ΔΔCt). The hsa-miRNA primer was: miR-210 5' CUGUGCGUGUGACAGCGGCUGA 3'.
HIF-1α small interfering RNA (siRNA) production and transfection
Single-stranded siRNA specific for human HIF-1α (si-HIF-1α) was synthesized by Bioneer (Korea). Sequence of the oligonucleotide was as follows: 5' AGAGGUGGAUAUGUGUGGGdTdT 3'. For transfection, hMSCs were grown to 80% confluence in 10% FBS contained DMEM at 37℃ incubator and then transfected with HIF-1α siRNA using Oligofectin (Invitrogen, USA). Briefly, si-HIF-1α was added to hMSCs at a final concentration of 100 nM in 10% FBS contained DMEM without antibiotics. After transfection for 4 h, the media were replaced in 10% FBS contained DMEM with antibiotics.
Western blotting analysis
hMSCs were lysed in lysis buffer containing phosphate-buffered saline containing 1% Triton X-100, 10× protease inhibitor cocktail (Roche, Switzerland), 1 mM phenylmethylsulfonyl fluoride (Thermo Fisher Scientific, USA), and 60 nM MG132 at 4℃ for 30 min. Protein concentrations were measured using the Bradford Protein Assay Kit (Bio-Rad, USA). The protein samples (60 µg) were then subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The separated proteins were transferred onto polyvinylidene fluoride membranes (EMD Millipore, USA) at 100 mV and 135 mA for 2 h by Power supply (Bio-Rad, USA). The membranes were blocked in Tris-buffered saline-0.1% Tween 20 (TBST) containing 10% nonfat dried milk for 1 h at room temperature or overnight (O/N) at 4℃. Next, the membranes were incubated with 1 : 1,000 anti-HIF-1α, 1:4,000 anti-β-actin, 1:1,000 anti-phospho-Akt, 1:1,000 Akt, 1:1,000 anti-phospho-ERK1/2, and 1:1,000 ERK1/2 antibodies for 1 h at room temperature or O/N at 4℃. The blots were washed in 1× TBST three times at room temperature and subsequently incubated with 1:4,000 horseradish peroxidase-conjugated secondary antibody against mouse or rabbit (Santa Cruz Biotechnology, USA) for 1 h at room temperature. After washing the membranes six times using TBST, immunoreactive proteins were detected using an enhanced chemiluminescence (ECL) system (Amersham Biosciences, UK). ImageJ software (National Institutes of Health, USA) was used for band intensity quantification.
Cell viability assay
Cell viability was determined with a CCK-8 [2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfo phenyl)-2H-tetrazolium, monosodium salt] assay kit (Dojindo, Japan). hMSCs (5 × 103 cells per well) were plated in triplicate wells in 96-well plates (Corning, USA) and incubated for 24 h. The cells were transfected with miR-210 mimic and incubated in a hypoxic chamber for 24 h. The CCK-8 assay kit was diluted with 0.1% FBS contained DMEM and then 100 µL was added to each well and incubated for 2 h at 37℃. The cells were incubated at 37℃ for 2 h. Absorbance was then measured at 450 nm with a spectrophotometer (Bio-Rad, USA).
Results
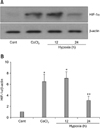
Expression of HIF-1α and miR-210 correlates with hypoxia and CoCl2 treatment
The activity of HIF-1α and miR-210 expression is increased by hypoxia. We first confirmed that HIF-1α was expressed in hMSCs under hypoxic conditions and with CoCl2 treatment using a Western blot. HIF-1α expression was up-regulated after CoCl2 treatment or with oxygen deprivation (Fig. 1). Until 24 h of hypoxia, HIF-1α expression increased compared to control. To detect miR-210 transcription, we examined changes in miR-210 transcript levels in the presence of CoCl2 or under hypoxic conditions. Changes in miR-210 transcription were the same as those observed for HIF-1α under the same experimental conditions (Fig. 2). Taken together, these data indicate that long-term exposure to hypoxia results in decreased HIF-1α and miR-210 expression.
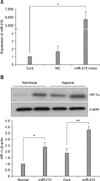
miR-210 positively regulates HIF-1α activity
To investigate the role of miR-210 in regulating HIF-1α expression, hMSCs were transfected with miR-210. Transfection efficiency was determined by real-time PCR (Fig. 3A). As shown in Fig. 3B, miR-210 overexpression induced HIF-1α expression under normoxic conditions and restored the expression of HIF-1α 24 h after hypoxia. These data suggested that an important function of miR-210 is to induce the protein expression of HIF-1α with normoxia and that this function is maintained during hypoxia.
miR-210 expression is strongly induced by HIF-1α
We investigated the link between miR-210 and HIF-1α up-regulation in hypoxic hMSCs. We successfully obliterated HIF-1α expression in hMSCs using siRNA. miR-210 overexpression enhanced HIF-1α protein levels in hypoxic hMSCs (Fig. 4). Furthermore, hypoxic conditions failed to induce miR-210 expression in the hMSCs with a concomitant loss of HIF-1α. Taken together, these results suggested a positive feedback loop between miR-210 and HIF-1α in hypoxic hMSCs.
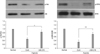
Survival of hypoxic hMSCs transfected with miR-210
To further evaluate the effect of miR-210 and HIF-1α on the survival of hypoxic hMSCs, we transfected hMSCs with miR-210 mimics and siRNA specific for HIF-1α. The cells were then maintained under hypoxic conditions. Survival was measured with a CCK-8 assay. After 24 h of incubation with hypoxia, cell death was reduced in the miR-210-transfected cells compared to the other groups (Fig. 5). We also observed greater phosphorylation of Akt and ERK1/2 in the miR-210-transfected hMSCs under hypoxic conditions. Moreover, survival rate was significantly reduced in hMSCs transfected with siRNA specific for HIF-1α (Fig. 6).
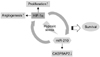
Based on our results, we proposed a model in which HIF-1α induces miR-210 expression that in turn increases the stability of HIF-1α. This positive feedback loop helps to increase HIF-1α levels during low-oxygen conditions and results in increased hMSC survival under hypoxic conditions through enhanced phosphorylation of Akt and ERK1/2 (Fig. 7).
Discussion
In the present study, we demonstrated the protective effect of miR-210 and HIF-1α in hypoxic hMSCs. Based on our findings, we suggest that a positive feedback loop involving these factors results in increased survival of hMSCs cultured under hypoxic conditions via Akt and ERK signaling. MSCs, which are multipotent stromal cells, have been isolated from various adult tissues such as born marrow, adipose tissue, muscle, and synovial membrane [24]. MSCs are capable of self-renewal and differentiation into multiple cell lineages [3,24,27]. Because of these capacities, MSCs have emerged as the most suitable candidates for use in cell-based therapy and tissue engineering. Despite interest in the clinical applications of MSCs, therapeutic techniques using these cells are limited because of poor MSC viability after cell transplantation. Thus, genetic modification to protect MSCs from apoptosis resulting from hypoxic conditions is necessary to improve the efficacy of MSC-based therapy.
miRNA is endogenous noncoding RNA that down-regulates gene expression by translational inhibition or degradation of target mRNA. The discovery of miRNA resulted in a paradigm shift in the way we view the role of RNA in regulating gene expression [1,7]. miRNA acts in a complex functional network in which each miRNA probably controls hundreds of distinct target genes and the expression of a single coding gene can be regulated by several miRNA molecules [20,22]. miRNA is made in a multi-step process. First, primary miRNA (pri-miRNA) transcripts are produced by RNA polymerase II in the nucleus. pri-miRNA is then transformed into hairpin precursors (pre-miRNA). The pre-miRNA is transported from the nucleus to cytoplasm where it is processed by Dicer to form mature single-stranded miRNA. Finally, a single strand is preferentially loaded into the core binding RNA-induced silencing complex. miRNA binds directly to the 3'-untranslated region of specific mRNA transcripts through complete or partial complementary pairing to induce silencing or direct cleavage of the mRNA. Recent evidence shows the important role of miRNA in regulating a multitude of physiological functions such as stem cell differentiation, neurogenesis, hematopoiesis, immune responses, skeletal and cardiac muscle development, and stress [4,9,11,16,17,26,33,36].
Induction of HIF-1α expression constitutes an integral part of the cellular response to hypoxia. Moreover, HIF-1α serves as a master regulator that orchestrates the expression of genes and helps cells adapt to low oxygen tension. HIF-1α translocates into the nucleus under hypoxic conditions to form a functional complex [30]. With a better understanding of gene regulation by transcription factors with the participation of miRNA, a novel relationship has been established between hypoxia and a select group of miRNA.
The expression of several miRNA transcripts has been found to be regulated by HIF-1α in response to hypoxia Among them, miR-210, a direct transcriptional target of HIF-1α, has emerged as a critical element of the cellular hypoxia response in a broad variety of cell types ranging from cancer cell lines to human umbilical vein endothelial cells [8,12,13]. Interestingly, miR-210 expression is regulated by both HIF-1α [2,5,12,35] and HIF-2α [8]. HIF-1α directly binds to a hypoxia responsive element (HRE) on the proximal miR-210 promoter [12]. When the miR-210 core promoter is compared across species, the HRE is highly conserved, indicating the importance of hypoxia in regulating miR-210 expression [13]. Consistent with this observation, the induction of miR-210 expression with hypoxia is dependent on HIF-1α [5]. miR-210 modulates hypoxia-induced cell cycle arrest [8,12,18]. Recently, miR-210-based gene therapy using mini genes in mice has been shown to improve cardiac function in a model of myocardial infarction [15]. Ischemic preconditioning also augments the survival of stem cells via miR-210 expression by targeting caspase 8-associated protein 2 [8].
Despite these findings, our understanding of the relationship between miR-210 and HIF-1α in hypoxic hMSCs remains limited. The purpose of the present study was to examine mechanisms underlying the induction of miR-210 expression via HIF-1α and evaluate the effect of miR-210 on the survival of hypoxic hMSCs. In conclusion, our results have helped increase our understanding about the relationship between HIF-1α and miR-210. Findings from the current study suggest that the viability of MSCs could be enhanced by increasing miR-210 expression before transplantation. Our data may serve as a basis for developing novel therapeutic modalities for treating injured tissues.



 XML Download
XML Download