

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Transmissible spongiform encephalopathies (TSEs) are a group of fatal neurodegenerative diseases caused by the accumulation of host-encoded prion protein (PrP) with an abnormal conformation mainly in the nervous system [33]. TSEs occur naturally in small ruminants. One example of is scrapie in sheep, which is the oldest known TSE first reported in the 18th century [9,26]. Unlike bovine spongiform encephalopathy (BSE), scrapie is not directly transmitted to humans [39]. However, scrapie prions are extremely resistant to degradation and can remain infectious for years in the natural environment [17].
In sheep, susceptibility to scrapie is determined by host genetic factors [18]. More precisely, this susceptibility is mainly modulated by amino acid polymorphisms encoded by the PrP gene (PRNP) at codons 136, 154, and 171 [1,3,12,23,24,38]. These polymorphisms are as follows: alanine (A) or valine (V) at codon 136, arginine (R) or histidine (H) at codon 154; and glutamine (Q), R or H at codon 171. Haplotypes for these three codons are commonly noted by three single-letter amino acid codes in the order of the codons (e.g., VRQ).
In Belgium, only the following five haplotypes are commonly present (listed in decreasing frequency): ARR, ARQ, ARH, AHQ, and VRQ [34]. The 15 most common genotypes have been classified into five groups of decreasing resistance (Fig. 1). The ARR/ARR genotype confers the highest resistance to the classical form of scrapie followed by the ARR heterozygotes except for ARR/VRQ [12,23]. The VRQ haplotype is associated with the greatest susceptibility [2].
In 1998, an atypical form of scrapie was discovered in Norway and named Nor98 [5]. Since then, this type of scrapie has been described in several European Union (EU) member states including Belgium [6,7,15]. It is not yet clear whether Nor98 is a transmissible disease in the field or whether it represents a sporadic disease. Genetic resistance to this specific form is not the same as that observed for classical scrapie. Sheep with the ARR/ARR genotype do not seem to have resistance to Nor98. Cases of Nor98, including those occurring in Belgium, have been described in animals with ARR/ARR genotypes [8,28]. However, ARR/ARR sheep are not the most susceptible to atypical scrapie since a polymorphism situated at a fourth codon [leucine (L) or phenylalanine (F) at codon 141] also affects susceptibility to this form of scrapie [32]. Most atypical scrapie cases have been detected in sheep with F at codon 141 (F141). It seems that the haplotype ARQ with F141 (noted as AF141RQ) is associated with greater susceptibility to atypical scrapie [28,35]. Similarly, the AL141HQ haplotype confers susceptibility to atypical scrapie [31]. In order to eradicate classical scrapie, the EU has promoted a breeding program since 2004 to select for ARR haplotypes and eliminate VRQ haplotypes from sheep populations [13]. It is known that selecting for scrapie-resistant sheep does not seem to have adverse effects on the production or health of the animals [13,40]. Lambs with Q/R171 alleles grow even faster than Q/Q171 lambs [14]. However, except for in the Netherlands [20] few data are available on the effectiveness of ARR haplotype selection as a function of time. This information is essential for assessing the current effect of the program as a whole. For this reason, the present study was carried out to evaluate the impact of the EU breeding scheme on the occurrence of resistant PRNP genotypes in Belgian sheep from 2006 to 2011. The effectiveness of selecting for the ARR/ARR genotypes and the influence of sheep breed on genotype frequencies were evaluated.
Materials and Methods
Samples included in this study had been sent to the National Reference Laboratory for routine genotyping analysis. These were either non-coagulated blood samples sent by the owners of living sheep to determine the genotype for future breeding purposes, or blood and brain samples supplied by an institution involved in TSE surveillance (e.g., a diagnostic laboratory or slaughterhouse). This second type of sample was from sheep slaughtered for human consumption or that had died from natural causes.
Commercial kits were used to isolate genomic DNA from blood (UltraClean BloodSpin; MO BIO Labs, USA) or brain (DNeasy; Qiagen, Germany) samples. Genotyping of codons 136, 154, and 171 was carried out with real-time (RT)-PCR (iCycler; Bio-Rad, USA) consisting of an activation of Uracil DNA-glycosylase at 50℃ (for 2 min), a DNA denaturation at 95℃ (8 min 30 sec) and 40 denaturation cycles at 95℃ (15 sec) and 60℃ (1 min). The codon 141 genotype was determined for a subset of the samples using the same RT-PCR technique but that was developed later for this codon. For each codon, the DNA was amplified with the reverse and forward sequence detection primers mentioned in Table 1 (Applied Biosystems, USA). The 136 primers were also used for codon 141 genotyping. For codon interpretation, two TaqMan MGB probes (three for codon 171 with three possible amino acids) were used with Universal PCR Master Mix without AmpErase Uracil.DNA glycosylase (Life Technologies, USA; Table 1).
For the brain samples that are often of low quality, genotyping data were confirmed by restriction fragment length polymorphism (RFLP) and electrophoresis on a denaturing gradient gel (DGGE). For these samples, the fragment with the codons was first amplified by PCR with two primers (P8: CAG GTT AAC GAT GGT GAA AAG CCA CAT AGG and P143: CTG GGA TTC TCT CTG GTA CT, Life Technologies): 95℃ for 10 min, 41 cycles of 94℃ for 1 min, 58℃ for 1 min, 72℃ for 1 min and then 72℃ for 7 min. The amplicons were then digested with BspHI (37℃ for 2 h) and the fragments were resolved on an acrylamide gradient gel containing formamide and urea as denaturants. The genotypes were determined by comparison with known samples from older analyses with RT-PCR.
The data set consisted of findings from 6 complete years (from January 2006 to December 2011). Genotypes for a total of 6,437 sheep were determined. Gender was known for 5924 of the sheep and the birth year was available for 1765 sheep. The analysis of codon 141 was limited to sheep carrying at least one ARQ haplotype that were still available in 2010 when the RT-PCR was developed for this codon in our lab. Statistical analyses were carried out with MyStat 12 (Systat Software, USA). Frequency changes between the two halves of the study period were verified by one-tailed 2 × 2 Chi-square tests (p-value < 0.05).
Results
In 2006, the ARR haplotype was found in 61.4% of the sheep population analyzed; this number increased to 79.3% in 2011 (Fig. 2). During the same time period, the ARQ haplotype frequency diminished from 21.1% to 12.9% (Fig. 2) along with the VRQ haplotype frequency (2.0% to 1.7%; one-tailed Chi-square, n = 12,874, χ2 = 227, p = 0.015). Likewise, the frequency of two rare homozygotic genotypes remained low (AHQ/AHQ: 0.23% to 0.29% and VRQ/VRQ: 0.08% to 0.10%). In brief, during these 6 years of selection the frequency of ARR/ARR sheep was multiplied by almost 1.7 (from 38.3% to 63.8%, one-tailed Chi-square test, n = 12,874, χ2 = 227, p < 0.000; Fig. 1). ARR/ARR was the only genotype to increase in frequency. Over the years, the proportion of males among the genotyped sheep fluctuated between 40.5% and 57.2% with an overall average of 48.7%.
Additionally, the sheep that were born more recently showed a higher ARR haplotype frequency (increasing from 35.1% to 83.1%; Fig. 3). Based on the birth year, the average age of the animals tested was similar throughout the 6 years of the study (average: 493 +/- 82 days). The amino acid encoded by codon 141 was identified for 238 sheep carrying the ARQ haplotype: 97.9% L/L (n = 233), 2.1% L/F (n = 5), and no F/F.
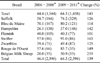
The breed was known for 5,163 (80.2%) of the sheep genotyped and 27 different breeds were analyzed. The majority of the animals belonged to nine breeds: Texel (the most common), Suffolk, Bleu du Maine, Hampshire, Flemish, Swifter, Zwartbles, Rouge de l'Ouest and Belgian Milk Sheep (Table 2). During the 6-year selection period, the number of ARR/ARR sheep increased for all the major breeds except among Flemish animals for which the ARR/ARR frequency remained about 40% (Table 3). These nine major breeds were roughly classified into three groups based on ARR/ARR genotype frequency and the frequency of genotype groups 3 or 5 (as defined in Fig. 1) as follows: first, breeds with the highest ARR/ARR frequencies but also the highest scrapie-sensitive ARR/VRQ frequencies (Swifter, Bleu du Maine, and to a lesser extent Rouge de l'Ouest; Table 4; second, breeds with average ARR/ARR frequencies (Suffolk, Texel, and Zwartbles); and finally breeds with the lowest ARR/ARR frequencies and highest ARQ/ARQ frequencies (Flemish, Belgian Milk Sheep, and Hampshire). The proportion of ARR haplotype significantly differed among these three groups (Chi-square test, n = 4,790, χ2 = 53.4, p < 0.000). The Belgian Milk Sheep breed was of further particular interest because these animals had a high proportion of ARR/AHQ.
Discussion
Over the 6-year study period (2006 to 2011), the selection program for increasing resistance against classical scrapie had a major impact on the ARR haplotype frequency in the Belgian sheep population. This was particularly evidenced by the increasing probability of carrying the ARR haplotype among more recently born sheep. Moreover, the already very low VRQ allele frequency was diminished even further. It is not clear whether the increased proportion of ARR/ARR sheep in the Belgian population has limited the number of (classical) scrapie cases. In Belgium, 78 scrapie cases have been detected in sheep since 1992 and 46 have been found since 2002 [41]. The last positive case occurred in May 2007 (i.e., during the first third of our study period). This relatively low prevalence and an absence of cases after 2007 limited the analysis of the impact of the breeding program on scrapie occurrence in Belgium. However, it is clear that there has been a decline of scrapie frequency in Belgium similar to that in other European countries [19,20,37].
Even if classical scrapie prevalence has diminished since the beginning of the breeding scheme, a number of Nor98 cases have been detected. Out of the 78 Belgian scrapie cases originating from 25 primary cases, nine (11.5%) were atypical (unpublished data). The first atypical case was detected in Belgium in 2002 [8]. Eight of the nine last scrapie cases in Belgium were atypical (detected between 2004 and 2007, unpublished data). This concentration of atypical scrapie in recent years cannot be attributed to changes in the age cohort of the sheep tested since the average age remained the same. Two atypical cases occurred in ARR/ARR sheep while none were found in VRQ/- sheep (unpublished data). However, ARR/- sheep are not more susceptible to atypical scrapie than other sheep [4]. The most common haplotypes with atypical scrapie are AF141RQ, AL141HQ, and AL141RQ [32]. In the limited number of samples analyzed, the AF141RQ haplotype (susceptible to atypical scrapie) has been found only very rarely [32]. This could indicate good prospects for increasing resistance to atypical scrapie in the current Belgian sheep population. Indeed, no atypical scrapie has been detected in Belgium for about 5 years.
The identification of atypical scrapie cases is probably due to better surveillance techniques. Yet it could perhaps be linked to the selection of sheep with genotypes that confer resistance to classical but not atypical scrapie. This represents a critical point for the EU breeding program. The occurrence of atypical cases in the sheep population under selection justifies continued monitoring for scrapie prevalence. Since BSE can infect sheep, this monitoring is also essential for understanding the theoretical risk of BSE in the sheep population [16,21].
In Belgium, sheep with the ARR haplotype represented about 70% of the sheep selected in 2008 and 79% in 2011. In the Netherlands, the percentage of ARR haplotype sheep also increased during the selection program, and in 2008 these animals accounted for about 55% of the sheep under active surveillance [20]. Among Baltic breeds not under selection, the ARR/ARR genotype was found in between 9% and 31% of the sheep under active surveillance in the Baltic states [36].
Even though the main factor influencing scrapie susceptibility is the actual frequencies of the different susceptible haplotypes, sheep breed can also have an effect [29]. This was confirmed in the present study as the breeds greatly varied in genotype frequencies. For example, Swifter sheep showed the genetic profile most highly resistant to scrapie (75% were ARR/ARR). Other studies have demonstrated that even within a given breed, ARR/ARR frequency before selection is highly variable. For example, this frequency ranged from 19% to 58% in Suffolk [10,11,22,25,27]. In Belgium, the Suffolk sheep tested in our study ultimately developed an ARR/ARR genotype frequency of more than 74%. Texel sheep are very common in Belgium (58% of the samples in this study). This breed had an average proportion of highly resistant sheep (44% being ARR/ARR at the beginning of the study) and was the only breed in this study that included almost all genotypes. Texel sheep represent good opportunities for further selection since 92% of all these sheep in our study had the ARR haplotype. Similarly, Texel is also the most common breed in the Netherlands. About 56% of the farms contain Texel sheep (and possibly other breeds) followed by Swifter (30%~34%) and then Zwartbles (12%~16%) [30]. At the end of our study in Belgium, 64%, 93%, and 49% of these three breeds, respectively, were found to have the ARR/ARR genotype. To our knowledge, no genotyping data according to breed is available for the Netherlands, but on the whole 32% of the sheep on farms adhering to a selection program have the ARR/ARR genotype [20]. In Germany, 11% of Texel sheep are ARR/ARR before selection [10]. For undetermined reasons, the ARR/ARR frequency did not increase among Flemish sheep during the breeding program in Belgium. In conclusion, our results indicate that the Belgian sheep population is relatively scrapie-resistant although this can still be improved.
It is important to control goats along with sheep populations for a global approach to eradicate scrapie. More information on scrapie genetics in goats is required before the feasibility and advantages of such a breeding program can be assessed. This matter is currently being evaluated by the EU [42].
In summary, the breeding scheme for Belgian sheep has been very efficient in terms of increasing the proportion of ARR haplotypes within only a few years. It is not yet clear, however, whether this has led to decreased classical scrapie prevalence in Belgium. Nevertheless, it is apparent that in terms of prevalence atypical scrapie has almost replaced classical scrapie in recent years (2002~2007). Since 2007, no cases of scrapie, whether classical or atypical, have been observed in Belgium. However, the current sheep population is still amenable to further selection even if variations between the breeds result in differences in selection possibilities. In particular, the Texel, the major breed in Belgium, is still a good candidate for selection given that only 54% of Texel sheep have the ARR/ARR genotype as well as almost all other gnotypes.






 XML Download
XML Download