

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Epizootic ulcerative syndrome (EUS) caused by Aphanomyces (A.) invadans is one of the most destructive diseases affecting several species of fresh and brackishwater fish both in farms and in the wild [10,20,28]. The key diagnostic features of EUS are the presence of the fungus and mycotic granulomatosis in the tissues of affected fishes [12,17,23,24]. The fungus is believed to be largely responsible for massive tissue damage and ulcers on the fish body. Ulcers and fish mortality are due to the invasive activity of the fungus [31]. However, not all ulcers in a fish are related to EUS because the lesions do not occur in epizootic proportions and are not seasonal in nature [26]. In addition, disease incidence and mass mortality due to EUS are reported to be promoted by several environmental conditions such as low temperature along with rapid change in salinity and dissolved oxygen [6,21,30].
Recently, EUS has re-emerged in endemic regions including Kerala (1999), Bangladesh (2001~2003), Nepal (2005), and Karnataka (2005) [1,5,7,18]. New occurrences of this disease are still being reported in previously unaffected areas and farming systems. During November 2006, EUS broke out in parts of the Zambezi River in Zambia and South Africa [19]. In 2009, snake head fingerlings with characteristic lesions collected from the wild in southern Florida (USA) were confirmed to be affected by EUS [25]. More recently, the first outbreaks of EUS were reported in Canada [27] and Australia during 2010 [3].
Traditionally, EUS is diagnosed by selectively growing the fungus from fish tissue in glucose peptone (GP) broth [11]. This method is not ideal for routine diagnosis. Histological examination, the most common technique used to diagnose EUS, is time-consuming [4]. Furthermore, in situ diagnostic methods such as in situ DNA hybridization [29], immunohistochemistry and immunofluroscence assays [9,16], are specific but time-consuming and require sophisticated equipment. Our group previously developed a sensitive monoclonal antibody-based immunodot assay that requires 3 h to complete [8]. Polymerase chain reaction (PCR) [20] is highly sensitive for EUS detection, but there are practical limitations to its widespread application due to its cost, ease of execution, need for trained personnel, and time required for completion. Development of field-friendly, rapid, sensitive, and low-cost tests will be highly useful for managing diseases such as EUS. The goal of the present study was to develop a simple and rapid monoclonal antibody-based flow-through immunoassay (FTA) for detecting A. invadans in fish.
Materials and Methods
Collection and preparation of fish samples
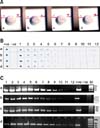
Fish with clinical signs of EUS (Fig. 1) and apparently healthy fish of the same species without ulcers were collected from a fish landing center in Mangalore, India. DNA from the fish was subjected to PCR to confirm infection with A. invadans [20]. Briefly, DNA from infected muscle was extracted using a KAPA Express Extract kit (Kapa Biosystems, USA). PCR was performed in 50-µL reactions using primers (forward primer: 5'-cttgtgctgagctcacactc-3'; reverse primer: 5'-acaccagatt acactatctc-3') developed by Oidtmann et al. [20]. Amplification was carried out in a thermal cycler (Bio-Rad Laboratories, India) with the following cycling parameters: initial denaturation at 96℃ for 5 min, 30 cycles of 96℃ for 1 min, 58℃ for 1 min, and 72℃ for 1 min followed by a final extension at 72℃ for 5 min. Purified A. invadans from culture was used as a positive control. PCR products were analyzed by 2% agarose gel electrophoresis.
Muscle tissue was excised from below or adjacent to the ulcerated area of the infected fish using a sterile scalpel and homogenized with 1 : 10 (w : v) TNE buffer (20 mM Tris base, 400mM NaCl and 50mM di-sodium EDTA, pH 7.4) in a 1.5-mL microfuge tube using a disposable microfuge pestle for 2~3 min. Muscle tissue from PCR-negative healthy fish was homogenized in a similar manner. The homogenates were allowed to settle for 5 min at room temperature and the cleared supernatant was used for the FTA.
Monoclonal antibody (MAb) against A. invadans
Hybridoma C14 producing MAb (IgM) specific for A. invadans was previously produced in our laboratory [9]. The hydridoma was grown in Roswell Park Memorial Institute (RPMI) medium for 4~5 days. The cell culture supernatant was used as MAb for the FTA.
Development of the FTA
The FTA was carried out according to Rajreddy et al. [22]. For this assay, a flow through cassette (Advanced Micro Devices, India) in which a nitrocellulose membrane (0.45 µm) is placed over absorbent pads to aid the flow of reagents (Fig. 2A). A. invadans isolate used in the present study was kindly provided by Dr. P.K. Pradhan (National Bureau of Fish Genetic Resources, Lucknow, India) and cultured as described by Lilley et al. [13]. A purified culture of A. invadans was used as the positive control and homogenized muscle from healthy fish (PCR-negative for A. invadans) served as the negative control. Positive and negative control samples were dotted on the upper side of the membrane above the control line. Homogenate from fish with EUS was dotted on the membrane below the control line. The samples on the membrane were air-dried for 5 min before the membrane was sequentially treated with 200 µL each of blocking buffer (5% bovine serum albumin, BSA in 50 mM PBS, pH 7.2; Sigma, USA), anti-A. invadans supernatant from the hybridoma (C-14), wash buffer (6% Tween 20 in 50mM PBS), Horseradish peroxidase (HRP)-conjugated rabbit anti-mouse IgG (1 : 500 in 50 mM Tris buffer saline, TBS-HCl and 0.2% BSA; Sigma), wash buffer, and a ready-to-use chromogen substrate TMB (3,3',5,5'-Tetramethylbenzidine)/H2O2 (Sigma) without any incubation periods in between exposure to each reagent. All the reagents were applied using plastic droppers (Fig. 2B). The reaction was terminated with 200 µL of wash buffer. Appearance of purple blue dots against the white background for the positive control and an absence of colour development for the negative control were recorded. Development of purple dots and their intensity for samples of the A. invadans-infected fish samples were compared to those of the positive and negative control. The entire FTA could be completed within 10 min. The assay was repeated several times to standardize the quantity and concentration of the reagents.
Comparing the sensitivity of the FTA with an immunodot assay using cultured A. invadans
A. invadans was cultured as previous described by Lilley et al. [13]. A. invadans mycelia were homogenized in TNE buffer (pH 7.4) and protein concentration of the homogenate was estimated according to the method by Lowry et al. [14]. The homogenate was subjected to serial double dilution. From each dilution, 2 µL were dotted onto nitrocellulose membranes for the FTA and immunodot assay. The FTA was performed as described above and the immunodot was performed according to the method by Gayathri et al. [8]. The lowest dilution of the homogenate that produced a purple dot in the immunodot and FTA was recorded.
Comparing sensitivity of the FTA, immunodot, and 1-step PCR using fish infected with A. invadans
Muscle tissues were collected from areas adjacent to ulcers from four different A. invadans-infected fish (Figs. 1A~D). The samples were prepared for the sensitivity test according to Anil et al. [2]. The muscle tissues from each fish were homogenized and divided into two equal aliquots. One part was added to TNE buffer (pH 7.4), homogenized, and subjected to log dilution. From each log dilution 2 µL were dotted onto nitrocellulose membranes for the FTA and immunodot assay. The two assays were performed as previously described. The lower limit of detection for A. invadans by the two methods was recorded. The other portion of the muscle tissue homogenates collected from the four fish (Figs. 1A~D) was subjected to DNA extraction. The extracted DNA was subjected to log dilution and PCR was performed for each log dilution.
Detection of A. invadans responsible for field outbreaks by FTA, immunodot, and PCR
A total of 73 fish with clinical signs of EUS from both cultured and wild populations were collected during the study period and brought to the laboratory on ice. From each fish, infected muscle was excised and divided into three equal parts for evaluation by FTA, immunodot, and PCR. Samples were prepared in respective buffers as described above and the final concentration was made up to 1 : 10 (w : v). FTA, immunodot, and PCR were carried out as previously described with appropriate positive and negative controls. Sensitivity of the FTA, immunodot, and PCR assay was calculated with the following formula [15,32]:

Stability of the FTA reagents
Two sets of FTA reagents were stored under different conditions: one at 4~8℃ and another at room temperature. FTAs were performed with A. invadans cultures using the reagents at 15-day intervals for 6 months. Shelf-life of the reagents was measured by evaluating the intensity of the purple dots that developed during the assay.
Results
Development of the FTA
Development of the FTA for analyzing homogenates
from infected fish along with positive and negative controls is described in Fig. 3. During the FTA, the positive control produced a blue-purple stain while the negative control did not. Positive samples yielded intense purple blue dots proportional to amount of A. invadans present in the samples (Fig. 3). The amount of reagents used for the assay was optimized to achieve consistent results.
Comparing sensitivity of the FTA, immunodot, and PCR assay using samples from fish with EUS ulcers
Muscle tissue homogenates from four fish with EUS ulcers (Figs. 1A~D) were diluted 10-1 to 10-12 with sample buffer. The samples were simultaneously evaluated with the FTA and immunodot. Similar log dilutions of DNA were evaluated by 1-step PCR. FTA and 1-step PCR detected A. invadans at a dilution of 10-11. In comparison, A. invadans was detected at a dilution of 10-8 by immunodot (Fig. 5). Overall, the FTA was as sensitive as 1-step PCR and 1,000 times more sensitive than the immunodot.
Detection of A. invadans in fish with EUS by the FTA, immunodot, and PCR
Muscle homogenate supernatants from 73 fish with EUS were evaluated for A. invadans with the FTA, immunodot, and PCR. The FTA and PCR assay detected A. invadans in all 73 samples (Table 2). The immunodot detected A. invadans in only 65 of the 73 samples (89.04%).
Discussion
In the present study, we developed an FTA for the rapid detection of A. invadans. We previously developed a similar assay for detecting white spot syndrome virus (WSSV) [22]. The FTA apparatus consisted of a nitrocellulose membrane placed on the top of adsorbent pads and enclosed in a plastic cassette. In the device, the adsorbent pads actively drew the liquid reagent through the membrane. The flow-through nitrocellulose membrane was capable of immunoconcentration to enhance the clarity and sensitivity of the assay. When commercially available, the FTA cassette will have pre-spotted positive and negative controls. Assay specificity was confirmed by reaction patterns of the positive and negative controls. Concentration of the blocking buffer, MAb, washing buffer, and enzyme-conjugated secondary antibody along with the reaction duration were optimized to achieve unambiguous results in 10 min. The assay is simple enough to use in the field.
The FTA detected A. invadans from pure cultures at a concentration of 7 µg/mL compared to 56 µg/mL by the immunodot. Increased sensitivity of the FTA compared to that of the immunodot is due to the immunoconcentration property of the FTA. Sensitivity of the FTA was also increased partly due to use of the sensitive chromogen TMB compared to 4-chloro 1-napthol used for the immunodot. Furthermore, sensitivity of the FTA was compared with that of the immunodot by using log dilutions of tissue samples from four fish with EUS. FTA detected A. invadans at a dilution of 10-11 compared to 10-8 by the immunodot. In a previous study, immunodot assays using MAbs C1 and C13 were found to detect 49 and 99 ng of fungus protein, respectively [9]. However, we used C14 due to the loss of C1 and C13 [9] that were previously used for immunodot assays to perform surveillance and predict EUS outbreaks in fish.
Sensitivity of the FTA was compared to that of 1-step PCR using log-diluted DNA extracted from tissue homogenates of four A. invadans-infected fish. Overall, FTA was found to be equally sensitive as 1-step PCR for detecting A. invadans in the infected fish sample. For the detection of white spot syndrome virus in shrimp, the FTA is 100 times more sensitive than 1-step PCR [22].
Finally, the ability of the FTA to detect A. invadans in fish samples with EUS ulcers collected during a field outbreak was compared to that of the immunodot and PCR assay. FTA and 1-step PCR detected the fungus in 100% (73/73) of the samples compared to 89.04% (65/73) identified by the immunodot. Results of this investigation further indicated that sensitivity of the FTA is equal to that of the 1-step PCR assay and is more sensitive than the immunodot. The immunodot could not detect fungus in all the fish with EUS symptoms because of its lower sensitivity. Additionally, six to eight fish samples can be tested in a row with a single FTA cassette. The FTA is therefore more suitable for use in the field compared to the immunodot and PCR.
Previously, the MAb-based immunodot was found to detect A. invadans in apparently healthy mullets 2 months before the appearance of EUS clinical signs [7]. The FTA developed in the present study is 1,000 times more sensitive than the immunodot and is simple to perform. Therefore, the FTA could be ideally used to routinely screen for A. invadans in aquarium fish at the port of entry and for EUS surveillance studies. A. invadans is spreading to new geographical areas [2,19,25,27]. In these regions, the FTA can be quite handy for specific detection of A. invadans and to differentiate EUS from other ulcerative diseases of less severity such as ones due to Aeromonas hydrophila.
FTA reagents stored at room temperature for 15 days or 4℃ to 8℃ for 4 months provided clear results. A similar study conducted by Rajreddy et al. [22] with RapiDot, an FTA used to detect WSSV, demonstrated that the assay reagents were stable up to 6 months when stored at 4℃ to 8℃ and for 1 month when maintained at room temperature. Differences in reagent stability may be due to the variations in characteristics of the MAbs used for the studies. MAb C14 used for the current study is an IgM that is less stable than IgG.
In summary, we found that the cost, sensitivity, and time required for performance are important features of effective screening assays. The FTA we developed was easy to use, cost-effective, and highly sensitive. Thus, this assay may be suitable for the rapid diagnosis of fish infected with A. invadans in the field during EUS outbreaks and at the port of entry during transportation.





 XML Download
XML Download