

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
There are two types of Peyer's patches (PPs) in calf intestine: ileal and jejunal [19]. These tissues consist of lymphatic follicles, interfollicular T cell areas, and dome regions [4]. The dome regions are covered by follicle-associated epithelium (FAE) containing "microfold" or "membranous" (M) cells which differ from intestinal epithelial cells morphologically and functionally [18,19].
The jejunal PP is thought to be a secondary lymphoid tissue similar to other mammalian PPs, and the FAE consists of M cells scattered among intestinal epithelial cells. In contrast, the ileal PP is thought to be composed of primary lymphoid tissue containing B cells and most areas of the FAE are covered by M cells [19,22]. M cells are important for initiating mucosal immune responses by transporting antigens into the underlying lymphoid tissues. These cells can also form entry points for pathogens such as invasive Salmonella Typhimurium which selectively penetrates and destroys M cells in intestinal PPs [15]. M cells may also deliver human immunodeficiency virus type 1 to target cells in mucosal lymphoid tissues [1]. In calves and lambs, prion protein has been found in FAE of the ileal PP, suggesting that M cells are important for uptake and therefore contribute to oral infection susceptibility [10,11,27].
Ulex europaeus agglutinin 1 (UEA-1) is a specific marker of mouse M cells [6]. CCL20 is a chemokine expressed in both mouse and human FAE [2,26,28]. Gebert et al. [8,9] have also shown that cytokeratin 18 and vimentin are useful markers of porcine and rabbit M cells, respectively. However, a bovine M cell-specific marker has not yet been identified. In addition, ontogeny and differentiation mechanisms of M cells are not fully understood and need further detailed investigation. In the present study, a monoclonal antibody specific for the FAE of calf ileal PPs was generated during ontogeny and characterized using immunohistochemistry, scanning electron microscopy (SEM), and Western blotting.
Materials and Methods
Animals
Japanese black calves (fetuses 5~9 months old, calves aged 2~10 days, 1~3 months, and 8 months; n = 3 for each developmental stage) were obtained from a local farm and slaughterhouses in the Miyazaki Prefecture (Japan). Fetal age was estimated by crown-to-rump measurements. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Miyazaki (Japan). The small intestines including ileal and jejunal PPs were collected for immunohistochemistry, SEM, and Western blotting. For immunohistochemistry, the specimens were mounted in an optimal cutting temperature embedding compound (OCT compound; Sakura Finetek Japan, Japan) on Cryomold (Sakura Finetek Japan, Japan), frozen on dry ice, and then stored at -80℃ until analysis.
Antibody production
Culture supernatant of a C6 hybridoma (C6) was generated as described previously [23] and used as a primary antibody. Briefly, mice were repeatedly immunized with a mixture of ovine monocyte-derived dendritic cells (DCs) generated in vitro and a population of ovine afferent intestinal lymphatic cells which contained approximately 15% mature DCs. Mouse antibody responses of CD11c+ ovine afferent lymphatic cells to DC surface antigens were assessed by a FACS Caliber flow cytometry (Applied Biosystems, USA) to identify the animal with the greatest response which then was subjected to a final boost consisting of only DCs. Screening of the hybridoma supernatants for DC binding was also performed by flow cytometry using afferent intestinal lymph cells [23].
Immunohistochemistry
Cryostat sections were stained with C6 using an indirect immunoperoxidase technique previously described by Yasuda et al. [30]. Briefly, sections (7- to 10-µm thick) were air dried on slides (Matsunami Glass Ind., Japan) and fixed with ice-cold acetone for 10 min. To block nonspecific binding, the sections were rehydrated in phosphate-buffered saline (PBS) and incubated with 10% normal goat serum (Vector Laboratories, USA) in PBS for 30 min at room temperature. The sections were incubated with C6 (culture supernatant of hybridoma) for 60 min at room temperature and washed three times with PBS. Next, the slides were incubated with biotin labeled horse anti mouse IgG (H+L) as a secondary antibody for 30 min at room temperature (Vector Laboratories, USA) absorbed with acetone powder of calf jejunal and ileal PPs. Endogenous peroxidase activity was then quenched with 0.3% H2O2 in methanol for 30 min at room temperature followed by incubation with ABC complex (Vector Laboratories, USA) for 15 min at room temperature. After the sections were rinsed three times in PBS, the reactions were visualized with metal-enhanced diaminobenzidine (Thermo Fisher Scientific, USA). Immunohistochemical staining was performed at room temperature in a incubation chamber (Cosmo-Bio, Japan). Control staining was simultaneously performed in which the primary antibody was replaced with normal mouse IgM (1,000 times dilution; Santa Cruz Biotechnology, USA). No positive staining was found in the control slides (data not shown).
SEM
Tissues were fixed in 2.5% glutaraldehyde (Nacalai Tesque, Japan), washed with 0.1 M phosphate buffer (PB; pH 7.4), and then further fixed with 1% osmium tetroxide (Merck, Germany) in 0.1 M PB. After dehydration with a series of graded ethanol solutions and substitution with isoamyl acetate, the specimens were dried in a critical point dryer (Type HCP-2; Hitachi, Japan) and coated with gold by ion sputter (E-1030; Hitachi, Japan). The specimens were then examined by SEM (S-4100; Hitachi, Japan).
Western blotting
Cells from calf ileal PPs were lysed in lysis buffer [TNE buffer (10 mM Tris-Cl, pH 7.8; 0.01% NaN3, 150 mM NaCl, 1 mM EDTA, and 1% NP-40), 1 mM phenylmethanesulfonylfluoride or phenylmethylsulfonyl fluoride, 0.3% aprotinin (Sigma-Aldrich, USA), and 0.1% phosphatase inhibitor cocktail 2 (Sigma-Aldrich, USA)], vortexed, and centrifuged at 15,000 × g for 20 min at 4℃. The proteins were heated to 96℃ for 5 min in Laemmli sample buffer (Bio-Rad Laboratories, USA), resolved on a 3~10% polyacrylamide gel in running buffer (Tris/glycine/SDS buffer; Bio-Rad Laboratories, USA), and transferred to polyvinylidene difluoride (PVDF) membranes (Immuno-Blot PVDF Membrane; Bio-Rad Laboratories, USA). The membranes were blocked overnight in blocking buffer (GE Healthcare Bio-Sciences, UK) at 4℃ and then incubated with C6 (culture supernatant of hybridoma) for 60 min at room temperature. The membranes were washed by 0.01% teen20-PBS, and incubated with secondary antibodies (horseradish peroxidase-conjugated anti-mouse IgM heavy chain, 100,000 times dilution; Millipore, USA) for 30 min at room temperature. Next, the membranes were again washed by 0.01% Tween 20-PBS and incubated with detection reagent (GE Healthcare Bio-Sciences, UK) for 5 min. A light capture system and CSanalyzer (ATTO, Japan) were used to detect antibody binding. Reactivity of C6 to the ovine antigens was also assessed using a standard procedure in which proteins from afferent intestinal lymph cells were lysed in 1% of NP-40, resolved on a 10% polyacrylamide gel, and visualized with a 3-amino-9-ethylcarbazole staining kit (Sigma-Aldrich, USA).
Results
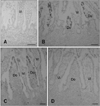
A summary of the C6+ cell distribution in calf PPs during ontogeny is shown in Table 1. During the middle fetal stage (Fig. 1A; fetal 5 months), no ileal PP lymphoid tissue formation or C6-positive reactions in the ileum were detected. During the late fetal stages (Figs. 1B and C; fetal 7 and 9 months, respectively), ileal PP formation was observed and C6+ cells were distributed in the FAE but not among the villous epithelial cells. Many cells covering the FAE were C6-positive and a small subset was C6-negative. Jejunal PP formation was also found during fetal development, but neither the FAE nor villous epithelial cells reacted with C6 (Fig. 1D; fetal 9 months).
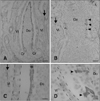
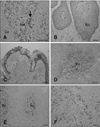
At the postnatal stage (2~10 days), almost the entire FAE in the ileal PP was C6-positive while the villous epithelium was negative (Fig. 2A). The epithelial cells changed from C6-negative to C6-positive along the crypt-to-dome axis (Fig. 2A) while all epithelial cells were negative along the crypt-to-villus axis (Fig. 2A). At the height of ileal PP development (around 1~3 months) and the start of involution (8 months), all epithelial areas in the FAE were C6-positive with a staining pattern similar to that found in PPs from the earlier postnatal stage (Fig. 2A). In addition, the cytoplasm as well as the membranes of cells in the FAE were C6-positive, but not in the villous epithelium (Fig. 2C). During the postnatal stages, very few positive cells were distributed in some FAE of the jejunal PP (Figs. 2B and D). The smooth muscles layer and smooth muscular fibers in the lamina propria were also C6-positive (arrows in Fig. 2). Hassall's corpuscle in the thymus medulla and epithelial layers of the palatine and pharyngeal tonsil were also C6-positive (Figs. 3A~C). In addition, reticular cells in the lymphatic follicle of the mesenteric lymph nodes, ileal PP lymphatic follicles, and the spleen were C6-positive (Figs. 3D~F). In these tissues, cells with the morphologies of DCs and macrophages also reacted with C6.
Morphological differences between M cells in FAE of ileal and jejunal PPs were observed by SEM. The villous epithelium of jejunal PPs had regular and dense microvilli during the postnatal stage (at 3 days) while M cells with irregular microvilli were sparsely scattered throughout the FAE (Figs. 4A and B). M cells with irregular microvilli covered the entire FAE of the ileal PP (Figs. 4C and D). In addition, cells similar to M cells in FAE of the ileal PP at the prenatal stage were also observed (data not shown). The monoclonal C6 antibody reacted with a bovine ileal PP protein as well as an ovine protein approximately 45 kDa in size expressed in migrating afferent intestinal lymph cells as shown by Western blot analysis (Fig. 5).
Discussion
FAE overlying lymphatic follicles of PPs in many domestic animals is a key player in the initiation of mucosal immune responses. Intermediate filaments such as vimentin, cytokeratin 8, and cytokeratin 18 have been identified as M cell markers in rabbit, rat, and pig, respectively [8,9,13,25]. In addition, lectin UEA-1 and peanut agglutinin are useful M cell markers in mouse and rabbit, respectively [6,14]. The chemokines CCL20 and CCL9 are expressed in mouse and human FAE. CCL20 recruits immature DCs to the FAE for antigen capturing and the induction of acquired immunity [2,26,28,31]. In calves, M cells can be distinguished from the villous epithelium by the different actin and villin staining patterns although these proteins are not exclusively expressed on the FAE [16].
In the present study, we described the C6 monoclonal antibody specific for a calf FAE antigen. We then analyzed staining patterns in the FAE of ileal and jejunal PPs during ontogeny. During the middle fetal developmental stage, jejunal but not ileal PP formation is observed [29]. At the late fetal developmental stage, C6+ cells were found in the FAE as described in this study. Previous histological observations indicated that the FAE is not developed in the ileum of calf fetuses at 73~150 days of gestation, but the FAE are distinguishable in fetuses 7~9 months old [3,5]. However, FAE in the jejunal PP did not react with C6 during the prenatal stages. According to the distribution of C6+ cells and the morphology of cells covering the FAE, M cells might therefore arise from epithelial cells in the ileal PP during the late fetal stage. In addition, the data suggest that FAE development in ileal PPs is different from that in jejunal PPs.
At the postnatal stages, C6+ cells covered all areas in the FAE of ileal PPs. In addition, very few C6+ cells were found in some FAE of the jejunal PP after birth. M cells were scattered throughout the FAE of jejunal PPs. Therefore, C6+ cell localization closely mirrored the distribution of M cells observed by SEM. However, additional studies are required to determine whether or not C6+ cells in the FAE of jejunal PPs are indeed M cells. Some DC and macrophages were also stained by C6. Furthermore, C6+ cells were found in the epithelium of Hassall's corpuscles of the thymus medulla and tonsils. We therefore speculate that the antigen recognized by C6 is a cellular protein that is involved in cell motility of antigen presenting, M, and epithelial cells. This hypothesis, however, requires further investigation.
Whether or not M cells are derived from epithelial cells is still unclear. It has been reported that human Caco-2 cells are converted into M cell by coming into contact with PP lymphocytes in vitro [17]. In addition, the absence of mature lymphocytes does not prevent the formation of M cells in the small intestine of Rag-1-/- mice in vivo [7]. M cells also develop in mouse intestinal villi [12]. However, the development of M cells in the villi is independent of the presence of lymphocytes while M cell development in the FAE is dependent on B cells [21]. This indicates that the signals for M cell differentiation vary between different anatomical compartments. In addition, previous studies of B cell-depleted fetal sheep showed that ileal PPs contain differentiated FAE and reticular cells [20,24]. Therefore, we speculate that development of the FAE in primary lymphoid organs is distinct from that of FAE in secondary lymphoid organs.
In conclusion, the results of our study showed that C6 is a useful marker of FAE in calf ileal PPs. C6 can also be potentially used for analyzing the functions and ontogeny of calf M cells in the FAE of ileal PPs.



 XML Download
XML Download