

 PDF
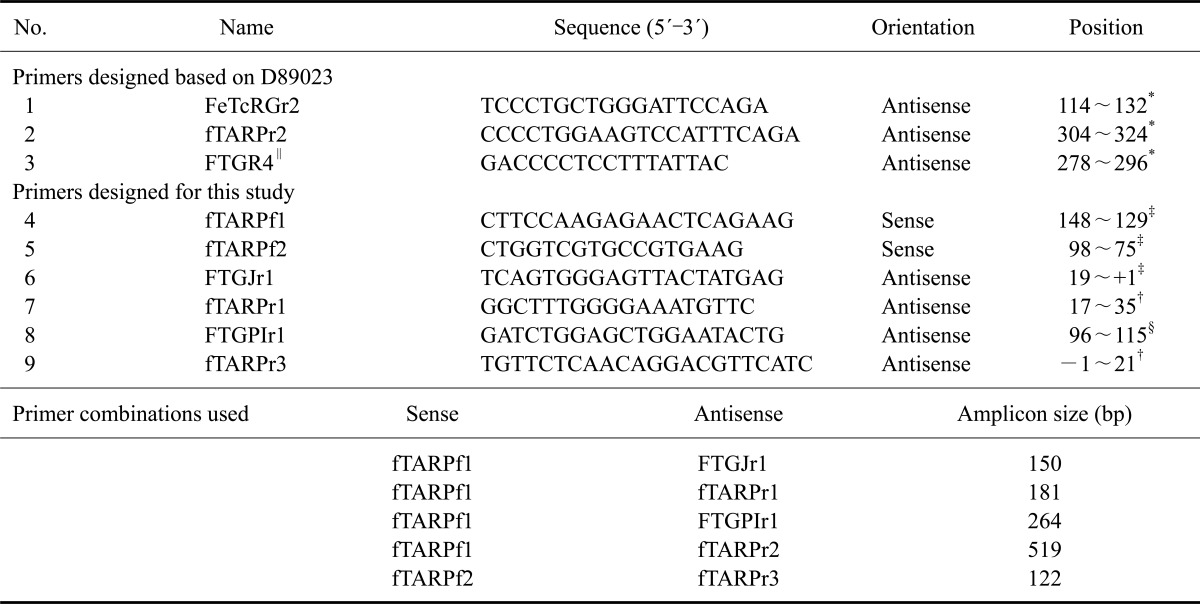
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The T-cell receptor γ alternate reading frame protein (TARP) is transcribed and expressed by epithelial cells of the prostate gland as well as benign prostate gland tumors, prostate cancer cells, and breast cancer cells in humans [8,9,30]. The transcript is derived from the T-cell receptor γ (TRG) J-region gene, TRGJP in germline configuration, and the constant region gene, TRGC1, of the human TRG locus. This region contains two open reading frames (ORFs) [8] but only the first is translated. The product is a 7-kDa protein with five leucine residues in heptad repeats followed by a basic region forming a potential leucine zipper motif [30]. The protein is located in the outer mitochondrial membrane, and its expression is increased in prostate and breast cancer cells [9,14,30]. Its expression in cancer cells in vitro leads to increased cell growth [29].
Because of TARP expression in prostate and breast cancer, this protein has been tested as a target for cytotoxic T-cells [1,7,19] and helper T-cells [12] as an innovative approach for specific adoptive tumor immunotherapy. Similar truncated transcripts originating from T-cell receptor α genes have been found in human and murine sarcomas. These proteins also seem to be involved in regulating tumor cell proliferation [6]. The TARP promoter has also been used to study adoptive tumor therapy [2-4].
TARP is transcribed from a J-region sequence and C-region sequence in germline configuration. In contrast, normal T-cell receptors are transcribed and translated from somatically rearranged genes of the same locus. This rearrangement of antigen receptor genes is known as V(D)J recombination [18]. Germline transcription of V-region and J-C-region genes takes place during V(D)J recombination within the thymus and their transcripts participate in the regulation of this process [13].
V(D)J recombination is initiated by products of the recombination activating genes 1 and 2 that bind to the recombination signal sequence (RSS) and induce DNA double-strand breaks [15,24]. The signal ends and coding ends then undergo non-homologous end joining [10]. The RSSs are composed of a highly conserved heptamere, a conserved adenine rich nonamere, and a less conserved spacer region of 12 ± 1 or 23 ± 1 bp. The length of the spacer is used to characterize the RSSs, which are annotated as 12RSS and 23RSS, respectively [10,22,23].
In the present study, we identified a feline transcript similar to human TARP composed of a J-region sequence and C-region sequence. This transcript was originally obtained from a splenic cDNA library established to characterize feline TRG sequences. Additionally, transcription of this construct was found in feline mammary neoplasm, similar to humans.
Materials and Methods
Animals and nucleic acid extraction
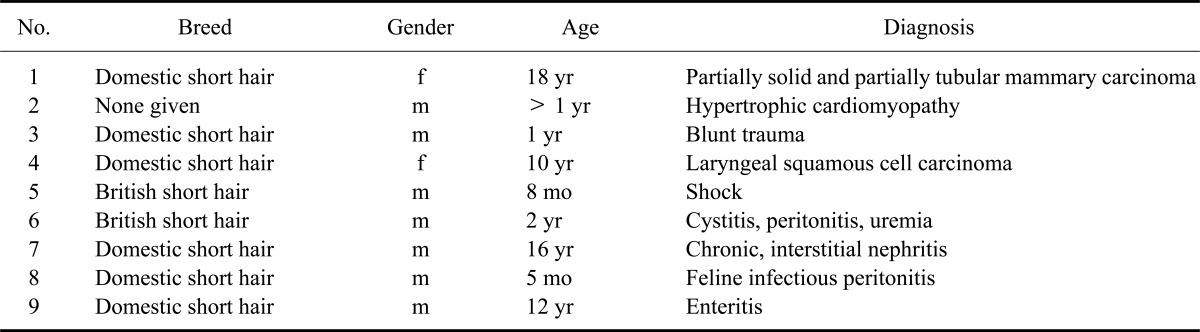
Total RNA was extracted from the spleens of three domestic shorthair cats of different ages and genders (see Table 1, cats Nos. 1~3) and the prostate gland of five male cats (cats Nos. 5~9) submitted for routine pathological evaluation to our departments. Prior to performing rapid amplification of cDNA ends (RACE), total RNA was extracted with a Purescript RNA Isolation Kit (Biozym Scientific, Germany). Genomic DNA was isolated from the same tissues using a Puregene DNA Isolation Kit (Biozym Scientific, Germany). For RT-PCR amplification of presumed feline TARP sequences from feline prostate glands, total RNA was extracted with a High Pure RNA Tissue Kit (F. Hoffmann-La Roche, Germany).
Nucleic acid extraction from formalin-fixed and paraffin-embedded (FFPE) tissues
To assess TARP transcription in feline mammary tumors, 32 FFPE neoplastic and 14 normal or hyperplastic mammary tissue samples from 22 female animals submitted for routine diagnostics procedures in 2008 and 2009 were analyzed. Samples of all major tissues from three additional animals excluding the spleen were pooled to evaluate transcription in other tissues. Total RNA was extracted from 30 to 40 paraffin sections of 2 µm thickness each. This was approximately equal to 10 mg of deparaffinized tissue. Tissue sections were deparaffinized by incubating with 800 µL xylene for 5 min at room temperature and pelleting at 14,000 × g at room temperature for 5 min. The resulting pellets were resuspended 300 µL of 99% ethanol and washed twice with 300 µL of 99% ethanol. Proteins were digested by resuspending the pellets in 300 µL cell lysis solution (Gentra Puregene Tissue Kit; Qiagen, Germany) and homogenizing by pipetting up and down. Next, 1.5 µL proteinase K solution (Gentra Puregene Tissue Kit; Qiagen, Germany) was added and the solution was incubated at 55℃ for 1 to 3 h. The samples were then inactivated by incubating at 94℃ for 10 min.
After cooling to room temperature, 800 µL TRIzol Reagent (Invitrogen, USA) were added and the samples were incubated at room temperature for 5 min. Next, 200 µL of chloroform (Carl Roth, Germany) were added. The samples were then incubated at room temperature for 5 min and centrifuged at 14,000 × g for 5 min at 4℃.
To remove residual phenol from the samples, an additional 400 µL of chloroform were added to the upper phase. After phase separation, 600 µL of isopropanol (Carl Roth, Germany) were added to the upper phase and samples were pelleted at 14,000 × g and 4℃ for 5 min. The pellets washed twice with 500 µL of 75% ethanol (Carl Roth, Germany), air dried, and resuspended in 20 µL diethylpyrocarbonate-treated water (Carl Roth, Germany).
5'RACE for feline TRG
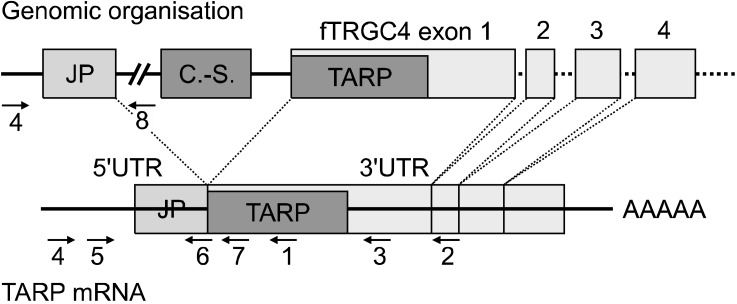
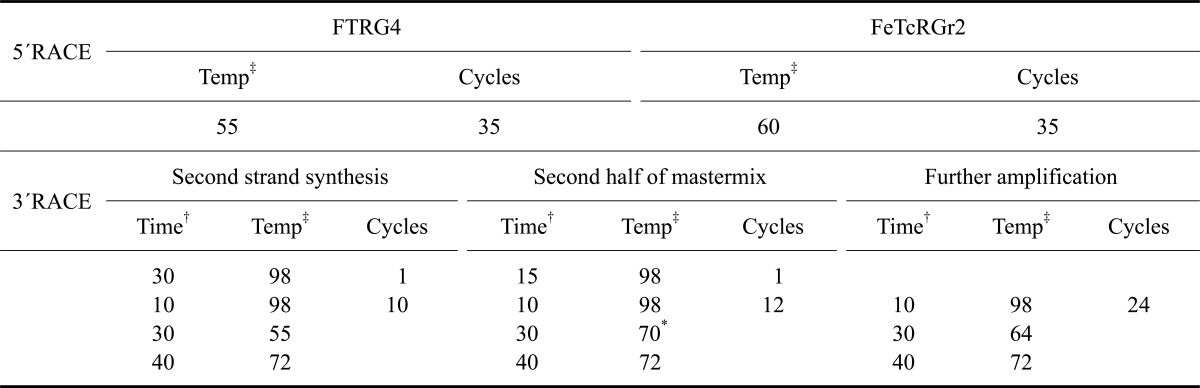
5'RACE was performed using a 5'/3' RACE Kit (F. Hoffmann-La Roche, Germany) and a Multicycler PTC 200 Gradient (Biozym, Germany). Amplification was carried out by nested PCR using Phusion High-Fidelity DNA Polymerase (BioCat, Germany) to ensure high sequence accuracy. The total reaction volume was 50 µL containing 1.5 mM MgCl2, 0.2 mM of each dNTP (Invitrogen, USA), 0.2 µM of each primer, 0.02 U/µL polymerase and 1 µg RNA. Primers were designed according to previously published sequences (D89023) [5]. FeTcRGr3 was used as a primer for cDNA synthesis, and FTGR4 along with FeTcRGr2 were used for amplification (for primer sequences, see Table 2). Primer annealing locations and the location of cloned sequences are shown in Fig. 1. DNA was denatured for 30 sec at 98℃, each cycle consisted of melting for 10 sec at 98℃, annealing for 30 sec, and elongation for 30 sec at 72℃ (for reaction conditions see Table 3). This was followed by a final elongation for 5 min at 72℃. PCR products were cloned with the AccepTor Vector Kit (Merck, Germany) and sequenced by a commercial laboratory (Eurofins MWG Operon, Germany) as previously described [26].
3'RACE of feline TARP sequences
3'RACE was performed using a 5'/3' RACE Kit (F. Hoffmann-La Roche, Germany) a Multicycler PTC 200 Gradient (Biozym, Germany). Amplification was catalyzed by Phusion High-Fidelity DNA Polymerase (BioCat, Germany) and performed in two steps. First, second-strand synthesis was conducted using the first half of a 50-µL PCR master mix composed as listed above containing 0.1 µM fTARPf1 as the gene-specific primer. After ten PCR cycles, the other half of the aforementioned master mix was added along with 0.24 µM of the second gene-specific primer fTARPf2 and 0.16 µM of the RACE anchor primer. Reaction conditions are listed in Table 3. The PCR products were cloned and sequenced as described above.
RT-PCR and PCR amplification of feline TARP
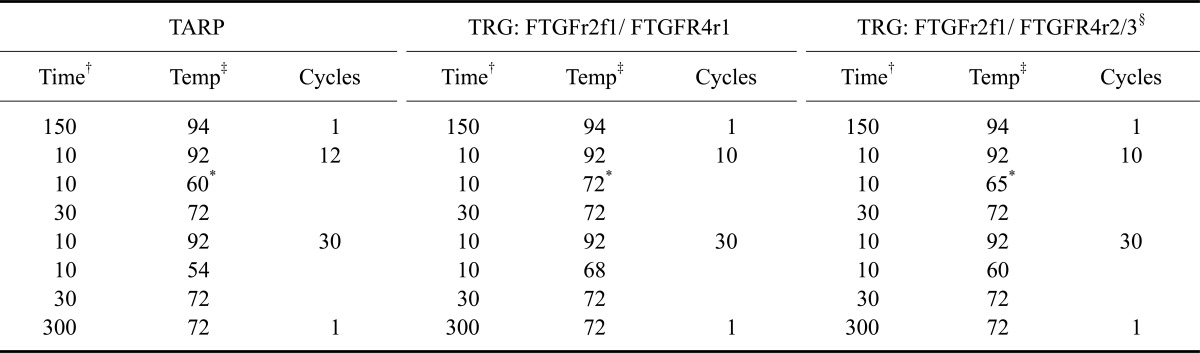
cDNA was synthesized using a RevertAid H Minus First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA) using the included random hexamer primers as recommended by the manufacturer's instructions. GoTaq DNA Polymerase (Promega, USA) was used to amplify cDNA and genomic DNA. The PCR reaction contained 100 ng DNA, 2.0 mM MgCl2, 0.2 mM of each dNTP, 0.2 µM of each primer, and 0.025 U/µL DNA polymerase. The final reaction volume was 25 µL and the PCR conditions are listed in Table 4. fTARPf1 and -2 were used as forward primers; FTGJr1, fTARPr1, fTARPr2, and FTGPIr1 were used as reverse primers.
Sequence analysis
Sequences were analyzed using the Basic Local Alignment Search Tool (National Center for Biotechnology Information, USA) and ClustalW (UCD Dublin, Ireland). GeneDoc 2.6.003 (Pittsburgh Supercomputing Center, USA) was used for displaying multiple sequence alignments. The sequences generated have been submitted to the Nucleotide Sequence Database (European Molecular Biology Laboratory, Germany): AM502837~AM502846, AM940997 (complete CDS), and FN433011.
RT-PCR amplification of TARP in feline mammary tumors
Approximately 500 ng of total RNA from each sample were reverse transcribed using RevertAid H Minus First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA). A PCR master mix of the same composition as described above was used. The fTARPf2 and fTARPr3 primers were used and the reaction conditions were slightly modified. Starting temperature for annealing was 63℃ followed by a touch down of 1℃ per cycle for 10 cylcles. Afterwards 57℃ were used. cDNA amplification was monitored using the YWHAZ housekeeping gene as previously described [20].
Results
Characterization of feline TARP
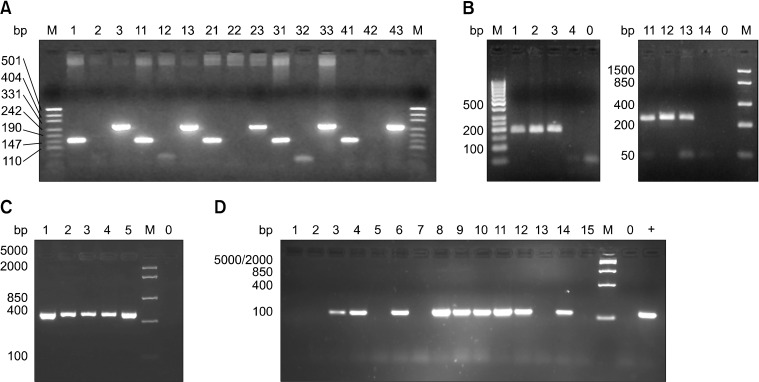
The seven clones (AM502839~AM502845) produced by a classical 5'RACE procedure using total RNA extracted from the spleens of two different cats (Nos. 2 and 3) included a transcript similar to human TARP (Fig. 2A). This transcript consisted of a TRG J-region sequence that was spliced to a C-region sequence (Fig. 1). To analyze the whole sequence of the transcript, 3'RACE was conducted with primers specific for the J-region sequence (cat No. 1). Resulting clones could only be sequenced in the forward direction. Sequencing with the reverse primer came to a halt within the poly-A-tail. Therefore, approximately 70 bp at the 3' end have not been sequenced. However, the sequence was almost identical (over 99% homology) to fTRGC4 that has been sequenced previously (GenBank accession No. AM489512). fTARP was also cloned and sequenced from total splenic RNA of cat No. 1 using fTARPf1 and FeTcRGr3/fTARPr2 (primers Nos. 4 and 2 shown in Fig. 1). This amplicon had more than 99% identity to the clones from animals Nos. 2 and 3.
To measure TARP transcription in the feline prostate gland, fTARP was amplified and sequenced from total prostatic RNA of five animals (Nos. 5 through 9; Fig. 3C) using the same primers. These amplicons also had more than a 99% identity to the sequenced clones. The C-region sequence encoded two ORFs. The first one started at the beginning of the C-region, contained 189 bp, and was out of frame compared to the fTRG transcripts. The second started at an 'ATG' site 185 bp downstream from the beginning of the C-region and contained 462 bp. This second ORF was in frame compared to the fTRG transcripts.
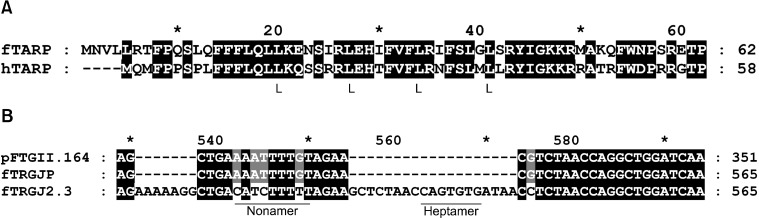
Sequence similarity of the coding sequence for human and feline TARP was 84% at the nucleotide level and 70% at the amino acid level (Fig. 2A). Sequence similarity of the second ORF between cats and humans was approximately 30% at the nucleotide level. The included J-region sequences had a 37% sequence similarity.
All feline C-region genes except for TARP/TRGC4 contained a start codon at the same position as human TARP. In TARP/TRGC4, a 'TTG' codon was found in this position. However, there was a start codon precisely at the beginning of the C-region. This position was located 12 bp upstream of the 'TTG' codon.
Similarity of the J-region sequence was 76% to fTRGJ2.1, 83% to fTRGJ2.2, 88% to fTRGJ2.3, and 80% to fTRGJ2.4. The J-region sequence in fTARP had a 19-bp deletion at the position of the 12RSS (Fig. 2B). This J-region was therefore named fTRGJP. The sequence, including a 5' intron, was also found in feline genomic sequences (gnl|ti|755161287) from the Trace Archive Nucleotide BLAST database (NCBI, USA).
To determine the genomic organization of feline TARP, the fTARPf1, FTGJr1, and fTARPr1 primers were designed. An amplicon recovered from genomic DNA using fTARPf1 and FTGJr1 was identified in all animals tested (Fig. 3A). The amplicon produced by fTARPr1, indicating the presence of spliced TARP DNA within the genome, was produced from the DNA of only one animal (No. 4; Fig. 3B). Additionally, the FTGPIr1 primer was specific for the intron downstream of fTRGJP. The amplicon produced by this primer was also recovered in all animals tested (Fig. 3A).
For cat No. 4, we amplified DNA pretreated with DNase and RNase using fTARPf1 and fTARPr1 or FTGPIr1. No product was generated from the DNase-treated DNA, indicating that both variants are present in genomic DNA (Fig. 3B). The amplicons generated by fTARPf1 and fTARPr1 or FTGPIr1 were cloned and sequenced. Similarity to fTARP was approximately 99% for both the genomic and cDNA sequences. Weak transcription of recombined TRG-V domains was detected within the prostate of cats Nos. 5~9 with the primers fTRGFR2f1 and fTRGFR4r1 (data not shown). Additional primer combinations yielded no amplicons.
Transcription of TAPR in feline mammary tumors
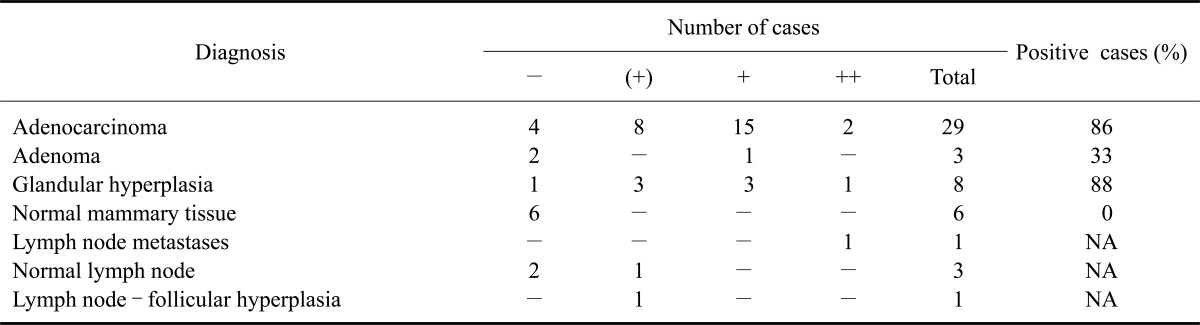
The amount of total RNA extracted from FFPE mammary tissues ranged from 0.6 to 46 µg (average: 8.6 µg, median: 5 µg, average 260/280 ratio: 1.6). Results of RT-PCR amplification of feline TARP sequences are summarized in Table 5. Because band density varied considerably, the results are presented in a semi-quantitative manner (Fig. 3D). The majority (81%) of the neoplastic tissues expressed TARP sequences whereas this expression was not observed in any of the normal mammary gland samples. No TARP mRNA was detected in the pooled RNA samples from other tissues of the three cats tested. All samples yielded bands for YWHAZ of approximately equal density.
Discussion
Transcription of a J-region sequence spliced to a C-region sequence is typical for human TARP. We detected the same constellation for one feline C-region and J-region sequence, suggesting that this transcript is the feline counterpart of human TARP. Whether these genes are located within the feline TRG locus, as is the case of human TARP [8,30], or comprise a separate locus that arose by gene duplication is not certain at the moment. Interestingly, the C-region sequence involved (fTRGC4) was never detected in combination with a functionally recombined variable domain [26-28]. Thus, we could not demonstrate its utilization as an fTRGC region. Furthermore, this C-region was associated with a carnivore-specific short interspersed element (CAN-SINE) located 468 bp in the 3' direction [27].
Due to its homology with rabbit TRG cDNA sequences (D38136, 86% nucleic acid identity), we first speculated that the presumed TARP we amplified could be a recombined VJ sequence. However, the sequence specific for the fTARPf1 primer is located within the intron upstream of TRGJP. An amplicon can thus be produced from genomic DNA using this primer and FTGJr1.
Interestingly, the start codon in this sequence was found 12 bp upstream from the corresponding position in human TARP. This indicates that after mutation of the original start codon present in all other feline C-region genes, a second mutation must have occurred. Therefore, fTRGC4 might have another role than that specific for a C-region of TRG.
The associated J-region sequence in fTARP features a 19-bp deletion within its RSS. fTRGJP is thus a pseudogene with regard to the TRG locus since productive recombination with a V-region is prevented. TR and immunoglobulin loci of all species tested so far contain multiple pseudogenes [10,13]. Pseudogenes arise from point mutations, deletions, or insertions after the regions have been duplicated. A pseudogene results if these mutations either alter the RSS or cause a reading frame shift [17,21].
Another possible manner in which the presumed fTARP transcript was formed, at least within splenic tissue, could be transcription of this sequence in a germline configuration. Each gene that encodes a region of the antigen receptors has its own promoter that is probably necessary for successful recombination [11]. These promoters induce transcription prior to recombination [10]. V-region and J-C-region genes are transcribed in germline configuration during recombination, but this takes place within the thymus in pre-T-cells and is induced by IL-7 signaling [13]. It is therefore unlikely that the alleged TARP transcript in spleen and prostate gland is caused by germline transcription. However, transcription of J-region genes in germline configuration has been observed in the spleen of fetal and adult cattle, but it has not been determined whether these transcripts could resemble TARP [16]. Excluding fTRGJP, we did not detect any transcription of fTRG sequences in germline configuration in feline spleen tissue. With regard to fTARP, this J-region sequence is only part of the 5' untranslated region.
Additionally, fTARP was amplified in cat No. 4 from genomic DNA in "spliced form". This finding indicates that in some animals fTARP additionally exists as a processed and reintegrated pseudogene. The fact that we found this pseudo gene in only one of the animals studied indicates that this transcript was very recently reintegrated into the cat's genome.
Similar to humans, alleged feline TARP transcripts were present in feline mammary adenocarcinomas and adenomas in the current study. However, cases of glandular hyperplasia were also positive for TARP transcription. This has not been reported in humans. Further testing should be conducted to determine whether TARP transcription indicates potential progression towards neoplastic transformation. Additionally, weak transcription found in lymphatic tissues should be further investigated because this might hamper the use of fTARP in tumor immunotherapy.
In summary, we have described an fTRC region highly transcribed in combination with an fTRGJ pseudogene in feline spleen and prostate as well as neoplastic tissues from feline mammary glands. The molecular characteristics were highly similar to those of human TARP. Further characterization of this transcript in cats could lead to a better understanding of its physiological function and significance. Additionally, if this mRNA sequence is also translated in feline cancer cells, it could serve as a tumor marker or a target for anti-cancer immunotherapy in this species. Furthermore, characterization of TARP in cats may result in a suitable animal model to analyze its potential as target for adoptive tumor immunotherapy.
 XML Download
XML Download