

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Brucella spp. are facultative intracellular Gram-negative bacteria and an important etiological agent that causes zoonotic disease. Brucella spp. are able to evade the host bactericidal phagocyte functions and proliferate within macrophages, thereby leading to the establishment of chronic infection [9,10]. Brucella spp. may occur as either smooth or rough, expressing smooth lipopolysaccharide (S-LPS) or rough LPS (R-LPS) as major surface antigen (Ag). S-LPS expressed by Brucella is the strongest Ag compared to other antigenic molecules that are involved in the immune response against brucellosis [16]. Currently, Brucella (B.) abortus S19 is used to immunize cattle whereas B. melitensis Rev 1 is used to immunize goats and sheep [28]. In general, the use of live attenuated organisms as vaccines is associated with safety concerns during vaccine production, and attenuated Brucella vaccines have many specific disadvantages, including abortion in animals administered during pregnancy [26,28]. For these reasons, different strategies are being sought for the production of safe, non-replicating vaccines that are easy to reproduce with consistent quality [20].
Recently, outer membrane proteins (OMPs) of Brucella have been evaluated as a non-LPS group of immunogens and vaccine [6,8,16]. Brucella OMP Ag are categorized according to their molecular weight into three groups: group 1, 2, and 3. Group 1, 2, and 3 Ags have approximate molecular masses of 94, 41 to 43, and 25 to 30 kDa, respectively [31]. All Brucella OMPs Ags, especially those in group 3, are also known as important factors that affect Brucella virulence [22]. These OMPs are major components of the sodium dodecyl sulfate (SDS)-insoluble cell wall fraction, and confer important vaccinal properties against Brucella infection [13,14]. The role of the two major members of OMPs, Omp25/Omp31 family, in protective immunity against Brucella infection is being studied by several groups [4,12,31]. Although Brucella OMPs Ags have important roles in immunogenicity and virulence, vaccines using OMPs Ags have not been fully evaluated.
We performed the present study to develop a subunit vaccine against Brucella infection in a mouse model. The gene encoding B. abortus Omp28 was cloned and expressed using a maltose fusion protein (pMAL) expression system. The ability of this recombinant protein (rOmp28) to protect against challenge with virulent B. abortus was evaluated along with the mouse response to immunization.
Materials and Methods
Bacterial strains and growth condition
A smooth virulent B. abortus biovar 1 strain of B. abortus 544 was kindly provided by Animal, Plant and Fisheries Quarantine and Inspection Agency in Korea and Escherichia (E.) coli DH5α cells was purchased from Invitrogen (USA). B. abortus was routinely cultured overnight in Brucella broth (BD Biosciences, USA) at 37℃ in a gyratory shaker (Waver Digital Platform; VWR International, USA) at 10 × g. When needed, solid medium was made by supplementing Brucella or Luria-Bertani (LB) broth (Becton Dickinson, USA) with 1.5% (w/v) agar (Takara, Japan). E. coli DH5α cells were used for producing the necessary plasmid constructs. E. coli cultures were routinely grown at 37℃ in LB broth or agar supplemented with 100 µg/mL of ampicillin (Sigma, USA).
rOMP expression
Total genomic DNA was prepared from B. abortus. Briefly, B. abortus cells were cultured at 37℃ overnight in Brucella broth with shaking. Next, 5 mL of the culture were collected and genomic DNA from the cells was recovered with a bacteria genomic DNA purification kit (iNtRON, Korea). The B. abortus Omp28 gene was amplified by PCR with the following primer pair: 5'-GATC GGA TCC AAC ACT CGT GCT AGC AAT TTT-3' (BamHI site underlined) and 5'-GATC AAG CTT TTA CTT GAT TTC AAA AAC GAC-3' (HindIII site underlined). The amplified DNA was digested with appropriate restriction enzymes (BamHI and HindIII; Takara, Japan) and ligated into a pMAL vector (New England Biolabs, USA). The recombinant plasmid was then used to transform the E. coli DH5α host cells. An exponential-phase culture of a rOmp28 clone confirmed in ampicillin-containing media was spread onto LB agar plates containing isopropyl β-D-1-thiogalactopyranoside (IPTG) (1 mM; Amresco, USA) and ampicillin (100 g/mL).
Purification of rOMP protein
One liter of LB broth supplemented with 100 µg/mL of ampicillin was inoculated with 10 mL of an overnight culture of bacteria expressing the fusion plasmid. IPTG was then added at a final concentration of 0.3 mM and the culture was further incubated at 37℃ for 2 h. Bacterial cells were harvested by centrifugation at 2,560 × g for 20 min. The supernatant was then discarded, and the cells were resuspended in 50 mL column buffer (20 mM Tris HCl, 200 mM NaCl, and 1 mM EDTA, pH 7.4). The sample was frozen at -70℃ and thawed in cold water three times. The cells were then was sonicated (Bandelin electronic, Germany) at 10,000 Hz in an ice-water bath and centrifuged at 5,200 × g for 30 min to collect the supernatant. The supernatant was diluted with column buffer (1 : 5) and loaded into maltose resin (Bio-Rad Laboratories, USA) according to the manufacturer's instructions. The purified protein was stored at -20℃.
Sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and immunoblotting
SDS-PAGE was performed according to previously described methods [11]. The purified rOmp28 was boiled for 10 min at 100℃ and diluted in 2× Laemmli sample buffer (4% SDS, 20% glycerol, 10% 2-mercaptoethanol, 0.004% bromophenol blue and 0.125 M Tris HCl, pH 6.8). After electrophoresis, the separated proteins were transferred onto Immobilon-P membranes (Milipore, USA) in transfer buffer (25 mM Tris, 192 mM glycine, and 20% methanol) with a constant current of 2 mA/cm2 for 30 min using a semi-dry electroblot assembly (Bio-Rad, USA). The membrane was blocked with 1% bovine serum albumin (Sigma, USA) for 1 h at 4℃, washed three times with 0.05% PBS-Tween 20, and incubated with Brucella-positive mouse sera (produced from mouse immunized by virulent B. abortus, 1 : 1,000 dilution) in blocking buffer at 4℃ for 2 h. The membrane was then washed with 0.05% PBS-Tween 20 and incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (1 : 10,000 dilution; Sigma, USA) in blocking buffer for 1 h at 4℃, and finally washed with 0.05% PBS-Tween 20. The proteins were detected with a luminol-coumaric acid-H2O2 detection solution (iNtRON, Korea) and exposure to X-ray film (Fuji, Japan).
Immunization and bacterial challenge in mice
Six-week-old female BALB/c mice (Japan SLC, Japan), two groups of five BALB/c mice each were immunized intraperitoneally with rOmp28 (100 µg/mL) or PBS in incomplete Freund's adjuvant (IFA) (Sigma, USA) on days 0, 14, and 35. Serum samples were obtained from all animals 14, 35, and 49 days after the first immunization via retro orbital bleeding. Serum titers of immunoglobulin G1 and IgG2a isotypes specific for rOmp28 were determined with an ELISA (BioTek, USA) according to the manufacturer's instructions. The cutoff value for the assay was calculated as the mean specific optical density for samples from nonimmunized mice diluted 1 : 100 as previously described [27].
To test the protective effects of rOmp28, mice were challenged with approximately 104 colony forming unit (CFU) of B. abortus 14 days after the last immunization. Ten days post-infection, all mice were scarified and the spleens were removed, weighed, and homogenized (Wisd homogenizer; Witeg, Germany) in saline. The homogenates were serially diluted (1 : 100~1,000) with PBS and plated on Brucella agar and then incubated for 3 days at 37℃. The number of CFUs for each spleen sample was determined to assess the virulence of the bacteria in each group of animals.
Results
PCR amplification and immunoreactivity of rOmp28
A specific band (753 bp) corresponding to B. abortus Omp28 was amplified by PCR. The PCR-amplified gene fragment encoding Omp28 was integrated into the pMAL expression vector and used to transform competent E. coli cells. The cloned product sequence was confirmed and found to completely match that of the reported Omp28 gene sequence for B. abortus (Genbank accession No. NC_007618.1). E. coli harboring the Omp28 plasmid were grown in LB medium, and 1 mM IPTG was used to induce Omp28 protein expression.
The protein product of the cloned Omp28 gene in the pMAL expression system was purified and was approximately 28 kDa (Fig. 1A). To evaluate rOmp28 protein immunoreactivity, Western blotting was performed (Fig. 1B). Our results showed that the purified rOmp28 strongly reacted with Brucella-positive mouse serum (1 : 1,000 dilution).
IgG1 and IgG2a responses to rOmp28 immunization
To test the protective effect of rOmp28, two groups of mice were immunized with purified rOmp28 protein or PBS with IFA on days 0, 14, and 35. Blood was collected from both groups 14, 35, and 49 days after the first immunization, and the IgG1 and IgG2a titers were determined. Throughout the entire experiment, the mean log titers of both IgG1 and IgG2a were significantly higher for the rOmp28-immunized mice compared to the animals injected with PBS (Figs. 2A and B). Our results clearly indicated that administration of rOmp28 increased both humoral and cell mediated immune response by inducing IgG1 and IgG2a production.
Protection of rOmp28 against B. abortus challenge
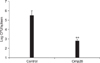
To analyze the ability of rOmp28 to protect against infection with virulent B. abortus, an in vivo protection study was performed in BALB/c mice (two groups with five mice each). In this experiment, protection was defined as a significant reduction (p < 0.05) in the mice receiving the vaccine when compared to animals injected with PBS. Mice treated with PBS developed splenomegaly that was enlarged 2~3 times its normal size as a consequence of the host inflammatory response, but this was not observed in the rOmp28-immunized group (Fig. 3). Vaccine efficacy was calculated by comparing the log units of bacterial burden in the spleens of the PBS-treated or rOmp28-immunized mice after B. abortus challenge. The mean log titers were 5.49 ± 0.49 for the PBS-treated animals and 2.89 ± 0.17 for the rOmp28-immunized (Fig. 4). These finding showed that immunization with rOmp28 conferred protection against challenge with virulent B. abortus.
Discussion
In a previous study, groups of Brucella OMPs were originally grouped according to their molecular weights and presences in outer membrane preparations [30]. OMPs play a vital role in maintaining the integrity and selective permeability of membranes [24]. In addition, expression of these proteins is often regulated by environmental signals; this characteristic is important for bacterial pathogenesis by enhancing adaptability to various environmental conditions [4,21].
Live attenuated Brucella vaccines interfere with serologic diagnosis of brucellosis because these vaccines induce a high-titer antibody response against the LPS O-polysaccharide. Antibody response is also limited to animals that develop an active case of brucellosis which is often delayed compared to a response against smooth LPS [19]. Accordingly, identification of Brucella protein components that can elicit an antibody response in the majority of infected animals would improve serological diagnostic methods and vaccinal strategies for treating brucellosis.
Development of a safe and effective vaccine against brucellosis has proven to be difficult. Indeed, numerous cell surface and intracellular components were examined as potential factors that could protect against Brucella infection. However, only low or intermediate levels of protection were achieved [1,5,29]. Regardless of the vaccine composition, a successful vaccination strategy would probably need to evoke a vigorous broad-based immune response. OMPs of Brucella spp. were characterized as immunogenic and protective Ags. Yet previous studies [4,7,12] were focused on the major OMPs (Omp25 and Omp31), and investigations on the protective capacity of minor OMPs were rarely performed. A previous report demonstrated that inoculation with B. abortus rOmp18 induces a Th1 response in vitro and the production of rOmp18-specific antibodies in BALB/c mice [30]. However, rOmp18 preparations were found to not protect against challenge with a virulent B. abortus strain [30]. Since Brucella species are facultative intracellular pathogens that reside mainly in macrophages, cellular immune responses are considered central for immunization [30].
Host immunity against B. abortus infection involves both Th1- and Th2-specific immune responses. Th1 responses are characterized by cellular immunity and the production of IgG2a antibodies. Th2 responses are characterized by humoral immunity, specifically the production of IgG1 [23]. The present study demonstrated that immunization with purified rOmp28 elicits protection against B. abortus infection in mice. Injection with rOmp28 produced in vivo Th1 and Th2 immune responses in the BALB/c mice (as shown by the production of IgG1 and IgG2a), and conferred a significant level of protection against challenge with virulent B. abortus 544. Because rOmp28 provided protection against B. abortus infection, this recombinant protein may be an important candidate for producing a vaccine against Brucella.
B. abortus infection results in a Th1 cellular immune response that promotes bacterial clearance [2]. Th1-isotype antibodies, such as IgG2a, are preferentially generated in humoral immune response by B cells against intracellular microorganisms, and may also opsonize the pathogen to facilitate phagocytosis [18]. During the course of infection, B. abortus is mainly cell-associated. However, this pathogen can be accessed through other mechanisms for clearance, such as ones mediated by IgG2a antibodies [15]. Presumably, these antibodies may have a greater ability than other isotypes to recognize microbial Ags on the surface of infected cells. The current study demonstrated that IgG2a and IgG1 responses were robust in B. abortus-infected mice vaccinated with rOmp28. It has been demonstrated that rOmp28 of B. melitensis can also protect against B. abortus infection [17]. In addition, rOmp28 of B. melitensis induced a humoral immune response in humans infected with both B. abortus and B. melitensis [22]. According to gene sequence data (Genbank accession no. NC_007618.1), the Omp28 homology between B. melitensis and B. abortus is almost 99% (NC_010742.1). These findings suggest that Omp28 is highly conserved among B. melitensis and B. abortus, and rOmp28 of both B. melitensis and B. abortus offers significant protection against virulent B. abortus infection.
The development of subcellular protein-based vaccines to prevent brucellosis has several advantages. One major advantage is the possible ability to differentiate vaccinated animals from infected animals. The current live attenuated vaccines against brucellosis for animals induce the high-titer production of antibodies against the O-polysaccharide of LPS, which interferes with ability to serologically diagnose the disease [3]. It has been reported that the B. abortus strain RB51, which is used for a general live attenuated vaccine, expresses the LPS O-side chain. Some cattle vaccinated with this strain were found to produce positive results for the card or buffered acid plate agglutination tests as well as the standard tube agglutination test [25].
In summary, the results from our study suggested that rOmp28 of B. abortus is a good vaccine candidate for protecting against B. abortus infection. However, the protective effects of rOmp28 in mice and the role of Omp28 in B. abortus virulence are still unclear. Consequently, further characterization of group 3 OMP genes of B. abortus including Omp28 will be required to understand the immunological and pathological aspects of brucellosis.



 XML Download
XML Download