

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The lack of estrogen is known to be a causative factor of osteoporosis [14]. It has therefore been suggested that estrogen is related to the differentiation and/or activities of bone cells, and many studies over the last few decades have investigated the relationships between bone metabolism and estrogen. Investigations have been performed in vivo using estrogen-injected [5,25] and ovariectomized animals [1,29], and the effects of estrogen on the differentiation and/or proliferation of osteoblasts and/or osteoclasts have been examined in vitro [11,22]. In addition, the expression of estrogen receptors (ERs) in bone cells has been evaluated both in vivo [7,30] and in vitro [17,26]. In the 1980s, a novel ER subtype, ER-β, was cloned in addition to the previously identified ER-α [9,31]. This discovery further complicated the relationship between bone metabolism and estrogen.
Formation and resorption of medullary bone in female birds is under the control of circulating estrogen [3,21]. Medullary bone can also be formed in the bone marrow cavity of male birds by estrogen administration [10,12]. ERs have been found in osteoblasts, bone-lining cells, and bone marrow cells in these experimental animal models [15,16]. As found in a previous study, ER-α mRNA, but not ER-β mRNA, is expressed at constant levels throughout the differentiation of osteogenic cells isolated from medullary bone [6]. Although these experiments were performed during the bone formation period, expression of ER-α and/or ER-β during bone resorption has not been assessed. In the present study, we analyzed the temporal expression of ER-α and ER-β mRNA during medullary bone formation and resorption in estrogen-treated male Japanese quails using reverse transcription-polymerase chain reaction (RT-PCR).
Materials and Methods
Animals
Male Japanese quails (Coturnix coturnix japonica; Quail Cosmos, Japan) 8~9 weeks old were used in our study. 17β-estradiol (E2) (Progynon-Depot, 20 mg/kg; Fuji Pharma, Japan) was injected into the breast muscle at 1, 2, 3, 5, 7, 10, and 15 days before the femurs were removed (n = 3~4). Control birds (day 0) were not treated with E2. All animal experiments were carried out in strict accordance with the Institutional Guidelines of the Committee of Research Facilities for Laboratory Animal Science, Hiroshima University, Japan.
Histological analysis
E2-treated and untreated (control) quails were sacrificed by decapitation and exsanguination. The right femurs were cut at the center of the diaphyses and fixed in 3.7% paraformaldehyde in phosphate buffered saline (PBS) (pH 7.4) at 4℃ for 5~7 days. The bone samples were then decalcified by immersion in PBS with 10% EDTA (pH 7.4) at 4℃ for 14~21 days. This solution was changed every 3 days. After decalcification, the proximal diaphyses were dehydrated in ascending grades of ethanol and embedded in plastic (Technovit 7100 kit; Heraeus Kulzer, Germany). Transverse sections of the diaphyses (2-µm thick) mounted on glass slides (Matsunami Glass Ind., Japan) were stained with hematoxylin and eosin.
RT-PCR analysis
The left femurs were quickly frozen in liquid nitrogen and stored at -80℃ until RNA was extracted. The diaphyses were crushed in liquid nitrogen and total RNA was obtained using a Sepasol RNA I Super commercial extraction kit (Nacalai Tesque, Japan). cDNA was synthesized from 3 µg of total RNA using a ReverTra Ace-α first-strand cDNA synthesis kit (Toyobo, Japan) and amplified with Taq DNA polymerase (Qiagen, Germany). Primers specific for ER-α (GenBank accession No. X03805), ER-β (GenBank accession No. AF045149), and β-actin (GenBank accession No. L08165) were designed using Primer 3 as a software for primer design (NCBI, USA). Table 1 summarizes the PCR amplification conditions with the specific primer sets. PCR was performed for 25 to 36 cycles. The amplified products were subjected to electrophoresis in 1.75% agarose gels and visualized with ethidium bromide staining. Band density was determined by densitometric analysis (ATTO Densitograph; ATTO, Japan). β-actin was used as an internal control.
Results
Histological analysis
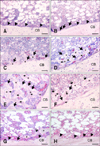
To examine histochemical changes in the bone marrow cavity following E2 treatment, transverse sections of femurs were stained with hematoxylin and eosin. In untreated male quails (day 0), the bone-lining cells appeared flat and were arranged on the endosteal bone surface. The bone marrow cavity was filled with bone marrow cells and adipose tissue (Fig. 1A). After E2 treatment, the bone-lining cells differentiated into cuboidal osteoblasts (day 1; Fig. 1B). These cells formed a part of the matrix between the endosteal surface and cells lining the bone (day 2; Fig. 1C). On day 3, matrices formed by the osteoblasts extended towards the bone marrow cavity and contained embedded osteocytes (Fig. 1D). These matrices were reticularly developed and many mature osteoblasts were seen on their surfaces on day 5 (Fig. 1E). Seven days after E2 treatment, volume of the matrices in the bone marrow cavity was reduced. This decrease was accompanied by an increase in osteoclasts and a decrease in osteoblasts on the surface of the matrices (Fig. 1F). After 10 days, the bone matrices were only observed in the vicinity of cortical bone while the marrow cavity began to refill with bone marrow cells and adipose tissue (Fig. 1G). After 15 days, the matrices had disappeared from the bone marrow cavity and the endosteal surface was again covered with flat bone-lining cells (Fig. 1H).
RT-PCR analysis of ER-α and ER-β expression
To assess the expression patterns of ER-α and ER-β mRNA during medullary bone formation and resorption, a semi-quantitative RT-PCR analysis was performed (Figs. 2 and 3). On day 0 (untreated quails), ER-α mRNA was already expressed but ER-β mRNA was barely detectable. The level of ER-α mRNA expression increased during bone formation although no significant difference was observed between days 0 and 5. Following this, ER-α mRNA expression decreased insignificantly from day 7 to day 15. ER-β mRNA was very weakly expressed throughout this period.
Discussion
In the present study, we measured the temporal expression of ER-α and ER-β mRNA during medullary bone formation and resorption in male Japanese quails treated with E2. Histochemical analysis demonstrated that medullary bone was formed in the bone marrow cavity by osteoblasts derived from bone-lining cells on the endosteal surface following E2 treatment. Many osteoclasts appeared on the surface of this bone 5 days after E2 administration. Following this, medullary bone disappeared from the bone marrow cavity and bone-lining cells reappeared on the endosteal surface after 15 days. This medullary bone model is therefore useful for examining the process of bone remodeling along with the relationship between estrogen and bone metabolism. This process of bone formation and resorption has not previously been reported in an experimental mammalian model. Several studies have previously investigated the relationship between mammalian osteoblasts and estrogen. Estrogen treatment stimulates cancellous bone formation in female rats [5]. Samuels et al. [24,25] demonstrated that the bone marrow cavity is filled with bone arising from cancellous bone in the proximal metaphysis after administering high doses of estrogen to mice. In addition, many studies using ovariectomized rats have reported the prevention of bone loss by estrogen [1,13,29].
In humans, ER-α and ER-β are expressed in cortical and cancellous bone, respectively, in cells such as osteoblasts, osteocytes, and osteoclasts [2,4,7,17,23]. Thus, the two ER isoforms may have different functions in different types of bone [4]. Batra et al. [2] demonstrated that the expression of ER-α and ER-β in human bone varied according to age, gender, and cell type. Oreffo et al. [17] found that ER-α mRNA is expressed in preosteoclasts but not mature osteoclasts. On the other hand, both ER-α and ER-β are expressed in osteoblasts, osteocytes, bone-lining cells, and osteoclasts on metaphyseal trabecular bone in rodents [4,7,32]. The maturation of osteoclasts found among human peripheral blood mononuclear cells [23] as well as murine bone marrow monocytes and RAW264.7 cells [26] is directly inhibited by estrogen. Additionally, the inhibition of bone resorption by estrogen via ER-α is mediated by a reduction of human osteoclastogenesis rather than by suppressing resorptive activity [27]. These results suggest that estrogen may affect bone formation and resorption through both ER-β and ER-α. Thus, it is not clear whether one or both types of ER is involved in the effect of estrogen on bone metabolism in humans and rodents.
The presence of ERs in avian medullary bone was previously examined during the early and active periods of medullary bone formation [15,16] using an ER-α-specific antibody [28]. Consequently, ERs were found in osteogenic cells such as bone-lining cells, osteoblasts, and alkaline phosphatase-positive bone marrow cells [16]. Using in situ hybridization, Imamura et al. [8] demonstrated that osteoblasts express ER-α mRNA but not ER-β mRNA. Furthermore, a previous study we performed indicated that osteogenic cells derived from medullary bone express ER-α mRNA, but not ER-β mRNA, during bone formation [6]. These results suggest that estrogen, acting through ER-α but not ER-β, might influence medullary bone formation by osteogenic cells.
Results of the RT-PCR analysis in the present study demonstrated that ER-β mRNA levels were very low while ER-α mRNA was stably expressed at higher levels during medullary bone formation and resorption. Although these results showed that expression of ER-α mRNA increased slightly, there was no significant change throughout the bone formation period. The variation in ER-α mRNA expression patterns might be due to differences between in vivo and in vitro studies, or could be related to the isolation of osteogenic cells during active medullary bone formation. Avian osteoclasts have been shown to express ERs [18,19], but Imamura et al. [8] recently demonstrated that osteoclasts do not express either ER-α or ER-β mRNA during the active medullary bone formation period. Although the presence of ERs in osteoclasts from medullary bone is debatable, the resorption activity of osteoclasts does seem to be inhibited by estrogen, suggesting that estrogen may regulate the expression of lysosomal genes [19] and the expression level of ERs in osteoclasts [20]. Reduced numbers of osteoclasts, as well as reduced osteoblast numbers and activity, might therefore be responsible for the slight decrease in ER-α expression we observed during bone resorption. As suggested by our RT-PCR analysis of total RNA extracted from the diaphyses of femurs containing cortical bone, medullary bone, bone cells, and other bone marrow cells, estrogen may act on osteoclasts via ER-α during medullary bone resorption.
In conclusion, our results showed that estrogen might affect both the formation and resorption of medullary bone through ER-α but not ER-β. Further studies are required to clarify which cell types, osteoblasts and/or osteoclasts, express ER-α during these processes. Moreover, we found that estrogen-induced medullary bone formation in male Japanese quails is a useful model for examining the relationship between bone metabolism and estrogen.



 XML Download
XML Download