

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Zearalenone (hexahydro-14, 16-dihydroxy-3-methyl-1H-2-deoxacyclotetradecine-1, 7(8H)-dione; ZEN) is a mycotoxin. This compound is a metabolite of fungi belonging to the genus Fusarium [1]. ZEN contaminates grains including barley, corn, oats, rice, and wheat or foods containing these grains [18,20]. Although ZEN has relatively low acute toxicity after oral administration to mice, rats, and guinea pigs, it produces endocrine effects, most importantly disruptions of the reproductive system, in animals [9,20]. ZEN is metabolized into zearalanol and zearalenol in animal tissues [6,12]. Its toxicity in animals depends on 3α-dehydroxylsteroid activity, which is involved in glucuronide conjugation and excretion of less toxic ZEN metabolites. Generally, carry-over of ZEN from contaminated feed to edible tissues such as meat, liver in pigs is negligible [7]. ZEN is considered to be a hepatotoxic, hematotoxic, immunotoxic, and genotoxic compound [20]. The maximum allowable concentrations of ZEN in food and animal feed have been established by many countries. The European Commission and other international governmental organizations have set maximum ZEN concentrations in parts per billion (ppb) for some foods and animal feed [7]. The United States does not have regulations pertaining to ZEN found in foods or feed, and there are no international action limits for ZEN despite the possibility of ZEN contamination of internationally traded cereal grains.
ZEN can be quantitatively analyzed using high-performance liquid chromatography (HPLC), gas chromatography-mass spectrometry, or ultra performance liquid chromatography-tandem mass spectrometry [10,13]. However, these methods require time-consuming extractions, sophisticated equipment, and skilled technicians. Thus, they are expensive to perform and not suitable for the routine screening of large numbers of samples in the field. Immunochemical techniques such as an immunochromatograpic assay [15], fluorescence polarization immunoassay [5], dipstick immunoassay [14] and enzyme-linked immunosorbent assay (ELISA) [1,16,19] are simpler and less expensive methods that have been developed for ZEN quantitation. Usefulness of these immunoassays is dependent on the specificity or sensitivity of the antibody used. In the present study, we produced a new anti-ZEN monoclonal antibody (mAb) with high specificity and affinity for natural ZEN, and then developed two assays: a direct competitive anti-ZEN antibody-coated ELISA and a direct competitive ZEN-coated ELISA.
Materials and Methods
Chemicals and reagents
ZEN, pyridine, carboxymethoxylamine (CMO) hemihydrochloride, dimethylformamide, N,N'-dicyclohexylcarbodiimide (DCC), casein, keyhole limpet hemocyanin (KLH), 8-azaguanine, hypoxanthine-aminopterin-thymidine (HAT) medium, Dulbecco's modified Eagle's medium (DMEM), bovine serum albumin (BSA), Tween 20, PEG 1500, Freund's complete adjuvant/incomplete adjuvant, and N-hydroxysuccinimide (NHS) were purchased from Sigma-Aldirch (USA). 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC) was purchased from Interchim (France). Goat anti-mouse IgG and 3, 3', 5, 5'-tetramethylbenzidine (TMB) were purchased from KPL (USA). All chemicals and organic solvents used were reagent grade or better. Monclonal antibody against ZEN was purchased from Santa Cruz (USA).
Experimental animals
Five female BALB/c mice (6 weeks old) were purchased from Orient Bio (Korea). The mice were given tap water and a commercial diet (Purina, Korea) ad libitum. The room housing the animals was maintained at a temperature of 24 ± 2℃, relative humidity of 50 ± 20%, and a 12-h light/dark cycle. All animals were cared for according to the Code of Laboratory Animal Welfare and Ethics of the Animal, Plant and Fisheries Quarantine and Inspection Agency (QIA) in Korea. The experimental design was approved by the QIA animal welfare committee.
Preparation of ZEN-oxime hapten
ZEN was first converted into ZEN-oxime to create a reactive group for coupling based on the method of Thouvenot and Morfin [17]. Ten milligrams of ZEN were dissolved in 2 mL pyridine, 20 mg CMO was added, and the mixture was stirred at room temperature (RT) for 24 h. The mixture was then dried with a hot plate stirrer (Corning, USA), and dissolved in 8 mL distilled water (pH 8.0). After being sonicated to suspend the residue, the aqueous suspension underwent three rounds of extraction with 3 mL benzene. Hapten was precipitated by the addition of 200 µL HCl and then extracted with 10 mL ethylacetate. The collected benzene phase was dried, dissolved in 8 mL distilled water and underwent another round of ethylacetate extraction. All ethylacetate phases were pooled, filtered over anhydrous sodium sulfate, and dried under a vacuum. The identity of the residue, ZEN-oxime, was validated by HPLC analysis using a mixture of water-acetonitrile (90 : 10, v/v) as the mobile phase. The number of haptenic groups per mole of BSA (epitope density) was calculated by dividing the moles of ZEN-oxime with those of BSA.
Preparation of ZEN protein and ZEN-enzyme conjugate
ZEN-oxime-BSA was synthesized using the carbodiimide condensation principle [2]. 1.64 mg of ZEN-oxime was dissolved in 0.2 mL dimethylformamide, mixed with 2.5 mL of phosphate buffer (0.15 M, pH 5.5) containing 5 mg BSA, and 25 mg EDC was added. The mixture was stirred for 16 h at RT. ZEN-oxime-KLH and ZEN-oxime-horseradish peroxidase (HRP) enzyme were synthesized according a previously published method [4]. Solutions of NHS (20 µL, 34 mg/mL in dioxane) and DCC (26 µL, 30 mg/mL in dioxane) were added to 2 mg of ZEN-oxime dissolved in dioxane (400 µL). The mixture was stirred overnight at RT. KLH and HRP (5 mg and 10 mg, respectively) were slowly dissolved in carbonate buffer (1.15 mL, pH 7.5) drop by drop and stirred at 4℃ for 6 h. After desalting with a PD-10 column (GE Healthcare, UK) according to the manufacturer's instructions, ZEN-conjugates were lyophilized and stored at -20℃ before use.
Production of mAb against ZEN
Five female BALB/c mice (6 weeks old) were acclimated for a week, and then immunized intraperitoneally with 100 µg of ZEN-BSA conjugate emulsified in an equal volume of Freund's complete adjuvant. Boosters with Freund's incomplete adjuvant were injected intraperitoneally 6, 8, and 10 weeks later. Four weeks after the last injection, serum was collected from each mouse and antibodies titers were measured with a ZEN-coated ELISA. Four days before spleen cell fusion with myeloma cell, a mouse that produced a high titer of antibodies with a greater affinity for ZEN was given an intraperitoneal injection of 100 µg ZEN-BSA conjugate without adjuvant. HAT medium selective to SP2/0-Ag14 (the American Type Culture Collection, USA) was prepared using DMEM containing 8-azaguanine. The ratio of spleen cells from the immunized mouse and fused SP2/0-Ag14 myeloma cells was about 5 : 1. After HAT selection, supernatant from the hybridoma cells was analyzed by a ZEN-coated ELISA. The selected cells were grown in 24-well plates and then cloned. A positive hybridoma cell was cloned by the limiting dilution method. The isotype of the immunoglobulin secreted from the cloned cell was determined using a mouse mAb isotyping kit (Roche, Switzerland). Immunoplates (Maxisorp; Nunc, Denmark) were coated overnight at 4℃ with 100 ng/well of ZEN-KLH conjugate in 0.05 M carbonate buffer (pH 9.6) and then washed three times with 0.05% Tween 20 in phosphate buffered saline, pH 7.0 (PBS-T). After blocking with PBS containing 1% casein (blocking buffer) for 2 h at RT, the plates were washed three times with PBS-T. A total of 100 µL of serum (or culture supernatant) were placed into the wells. Next, 100 µL of 500 ppb ZEN diluted in blocking buffer was added and the plates were incubated at RT for 1 h. After washing three times with PBS-T, 100 µL of anti-mouse IgG-HRP conjugate (1 : 2,000) was added. The plates were again incubated for 1 h at room temperature, and the washing steps with PBS-T were repeated. 100 µL of TMB substrate solution was added to the each well. After incubating for 15 min at RT, the reaction was stopped by adding of 2 N H2SO4 (100 µL per well). The absorbance was measured at 450 nm with spectrometer (Molecular Device, USA).
Production and purification of ascites containing mAb against ZEN
For ascites fluid production, a clone (kk-ZEN) which showing the highest affinity for ZEN was cultured in DMEM supplemented with 10% fetal bovine serum (DMEM-10). The cultured cells (2 × 106 cells) were washed three times with serum-free DMEM, suspended in 1 mL of DMEM and injected into mouse. After 7~14 days, ascites fluid was collected from the mouse and purified with a Hitrap protein IgG column (GE Healthcare, UK) according to the manufacturer's instruction. An indirect competitive ELISA was used to determine the presence of ZEN antibody in the purified ascites fluid.
Anti-ZEN antibody-coated ELISA
For the ZEN-coated ELISA, ZEN mAb (25 µg/mL, 100 µL per well) in 0.05 M carbonate buffer was immobilized on the immunoplate overnight at 4℃. After washing the plate with PBS-T, a ZEN standard or samples were diluted in 50 µL of PBS-MeOH buffer (containing 14% methanol) and 50 µL ZEN-HRP solution (1 : 300 in 1% casein), and added to each well. After incubating for 5 min at RT, the plates were washed with PBS-T and developed with 100 µL TMB solution for 5 min. To stop the color development, 100 µL of 2 N H2SO4 was added. The absorbance was measured at 450 nm in a spectrometer (Molecular Device, USA).
ZEN-coated ELISA
For the ZEN-coated ELISA, ZEN-KLH (1 µg/mL, 100 µL per well) in 0.05 M carbonate buffer was immobilized on the immunoplate overnight at 4℃. After washing the plate with PBS-T, a ZEN standard or samples were diluted in 50 µL of PBS-MeOH buffer (containing 14% methanol) and 50 µL of the anti-ZEN antibody-HRP conjugate solution (1 : 500 in 10% skim milk) were added simultaneously to each well. After incubating for 5 min at RT, the plates were washed with PBS-T and color development was performed as described above.
Measuring cross-reactivity of ZEN and its metabolites with both ELISA methods
Cross-reactivity of ZEN-related metabolites such as α-zearalenol (ZEL), β-ZEL, α-zearalanol (ZAL), and β-ZAL in both ELISA procedures was determined by dividing the 50% inhibitory concentration (IC50) of ZEN by the IC50 values of each metabolite.
Feed sample preparation
A total of 5 g of commercial pig feed (Purina, Korea) were spiked with different concentrations of ZEN (0, 150, 300, and 600 ng/g), and extracted once with 25 mL of 70% methanol/water (v/v) for 3 min by agitating. The extract samples were filtered through Whatman No.1 filter paper (Whatman, USA). A total of 10 mL of filtrate was diluted in 40 mL of PBS (pH 7.4). The concentration of ZEN in the extracted samples was determined using an ZEN antibody-coated ELISA described above.
HPLC analysis
Prior to analysis, extracted samples from feed were passed through an immunoaffinity column (R-Biopharm Rhone, UK) according to the manufacturer's instruction. For HPLC, a reverse-phase C18 column (3.9 × 150 mm, 5.0 µm; Waters, USA) was equilibrated with a water-acetonitrile solution (50 : 50, v/v) at a flow rate 1 mL/min. A total of 10 µL of each eluted sample was injected into an HPLC system (Waters 2695; Waters, USA) fitted with a fluorescence detector (Waters 2475; Waters, USA) set for 274 nm excitation and 440 nm emission wavelengths.
Results
Preparation of ZEN-protein/enzyme complexes as an immunogen
A ZEN-oxime peak was observed before 1 min and ZEN peak appeared around 8 min in HPLC chromatogram (Fig. 1). Formation of the conjugates was also confirmed by spectrophotometrical scanning between 190 and 340 nm. The UV spectra of ZEN-CMO included three maxima at 236, 271, and 302 nm; this was similar to those of ZEN alone that contained maxima at 235, 272, and 312 nm. These results indicate that modification with CMO did not affect the chromosphere characteristics of ZEN.
Production and characterization of ZEN-specific mAb
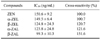
Nine cloned hybridoma cells were obtained and three produced the IgG1 subclass with κ-type light chains. We selected the clone that produced antibodies with the highest specificity for ZEN, which showed greater affinity and sensitivity than the commercially available ZEN antibody based on an indirect-competitive ELISA (Fig. 2). The clone was named kk-ZEN and used for mass production of mAb. Binding of the diluted mAb from kk-ZEN (250- and 500-fold) in microplates coated with 50 µg/well ZEN-KLH was inhibited by almost 0.1 ng/well of ZEN, but binding of the commercial anti-ZEN mAb was not effectively inhibited (Fig. 2). The IC50 values of the mAb for α-ZEL, β-ZEL, α-ZAL, and β-ZAL in the present study were 108.1, 119.3, 114.1, and 130.3%, respectively, based on the ZEN-coated ELISA and 100.7, 120.7, 121.6 and 151.6%, respectively, based on the ZEN antibody-coated ELISA (Tables 1 and 2).
Validation the method of ZEN- or anti-ZEN antibody-coated ELISA
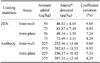
To obtain linear standard curves, we created a logit curve expressing the y-axis by calculating binding percentage using Microsoft Excel. Quantitative values for ZEN detection calculated from the ZEN antibody-coated ELISA and ZEN-coated ELISA results ranged from 25 to 750 ppb (r2 > 0.99) and from 12.5 to 100 ppb (r2 > 0.99), respectively (Fig. 3). The intra-plate and inter-well assay variations for both ELISA procedures were compared and expressed as a coefficient variation (CV). Two sets of standards with different ZEN concentrations (225 and 375 ppb for the anti-ZEN antibody-coated ELISA, and 50 and 70 ppb for the ZEN-coated ELISA) were analyzed. The intra- and inter-assay CV values were both less than 10%, indicating high reproducibility (Table 3). The standard curves for PBS buffered standards containing 5, 10, 14, 20, and 30% methanol had r2 values within same range despite the differences in ZEN detection between the 20 and 30% methanol assays (Fig. 4). Recoveries of spiked ZEN from feed ranged from 106.07 to 123.00%, and CV values ranged from 0.93 to 2.28% (Table 4).
Discussion
ZEN can react with specific antibodies but cannot induce the production of antibodies by itself. Therefore, a conjugate of ZEN and a carrier protein must be prepared for the production of ZEN-specific antibodies. After haptenization of ZEN with CMO, the chromatographic mobility of product (ZEN-oxime) appeared earlier than that of the parent compound as expected due to the increased polarity caused by acquisition of a reactive group [2]. In the present study, the ZEN-oxime peak also occurred before the ZEN peak and formation of the conjugates was also confirmed by spectrophotometrical scanning between 190 and 340 nm. The UV spectra of ZEN-oxime contained three maxima at 236, 271, and 302 nm, similar to those of ZEN alone that included maxima at 235, 272, and 312 nm. These results indicated that modification with CMO did not affect the chromosphere characteristics of ZEN. The number of moles of conjugated protein was calculated based on the concentration and molecular weight (BSA: 6.643 × 104 g/mol, KLH: 8 × 106 g/mol) of the carrier protein. The number of moles of ZEN-oxime was calculated based on the absorption at 316 nm using the extinction coefficient (ε = 6.02 × 103). The epitope densities of ZEN-oxime against BSA and KLH were reported differently depending on the experiment conditions. For the present study, a molar ratio of 1 : 25 was used. This ratio was slightly larger than the 1 : 19.8 ratio mentioned in another report [2]. Coupling density of the chemical with the carrier protein was reported to affect antibody production. Immunogens with higher coupling densities usually produce mAb with higher substrate affinities [8,11]. The immunogen used in the present study had a relatively high coupling density compared to those used in previous reports [2].
Antisera from the five immunized mice each showed different specificities and affinities based on an indirect competitive ELISA. The mouse that produced antisera with the highest affinity for ZEN was used for cell fusion with myeloma cells. Because the fused cells can produce many different antibodies, not only against ZEN but also against the BSA and CMO linkers, screening of the cells was performed by both indirect and indirect-competitive ELISAs using ZEN-KLH as the coating antigen. Nine cloned hybridoma cells were thus obtained and three produced an IgG1 subclass with κ-type light chains. We selected the clone that produced antibody with the highest specificity for ZEN, and had a greater affinity and sensitivity than a commercially available one based on the results of an indirect-competitive ELISA. Binding of the diluted supernatant to microplates coated with 50 µg/well ZEN-KLH was inhibited by 0.1 ng/well of ZEN, but binding of the commercial mAb was not effectively inhibited. These findings suggest that the supernatant from the kk-ZEN clone had a higher specificity for ZEN than the commercial anti-ZEN mAb.
The IC50 values of the mAb from the present study for α-ZEL, β-ZEL, α-ZAL, and β-ZAL were more than 100% based on the ZEN-coated and ZEN antibody-coated ELISAs. According to another report, the cross-reactivities of mAb produced in their laboratory were α-ZAL, β-ZAL, and β-ZEL were 42%, <1%, and <1%, respectively [3]. Another anti-ZEN mAb produced by author's laboratory also showed less than 1% cross-reactivity with these ZEN metabolites [2]. Based on the high affinity of the mAb developed in the present study for both ZEN and ZEN metabolites, it is likely that the antibody has a paratope that can bind to a structure common to both ZEN and its metabolites. Thus, this antibody could be used to simultaneously screen for ZEN and common ZEN metabolites in feed and food.
Even though both ELISA systems had a 10 ppb limit of detection and the intra- and inter-assay precision results were also satisfactory showing the below 10% CV values for both methods, our results indicated that the ZEN-coated ELISA had a narrower ZEN quantification range compared to the anti-ZEN antibody-coated ELISA.
Because ZEN can be extracted more easily from the solid matrices with organic solvents like methanol, we examined the effect of methanol on the ELISAs we performed by comparing standard curves obtained from PBS buffered standards containing methanol at concentrations ranging from 5 to 30%. Our results demonstrated that the ELISA system in this study was less affected by methanol than those described in previous studies [3,16], and that solutions containing up to 20% methanol can be effectively used as solvents for extracting ZEN from solid matrices without adversely affecting the assay results. Based on the Codex Alimentarius guidelines (CAC/GL 71-2009) for quantitative analytical methods, acceptable CV values for intra-laboratory testing are below 15% and acceptable recoveries for samples containing 102 to 103 ppb of the analyte are 70~110%. Recoveries in our study ranged from 106.07 to 123.00%, and CV values ranged from 0.93 to 2.28%, all which could sufficiently satisfy the standard regulations. Even though some nonspecific binding in samples spiked with ZEN into pig feed was observed, reproducibility was very satisfactory because CV values were below 3%.
In conclusion, we produced a very highly sensitive and group-specific mAb against ZEN and developed two direct competitive ELISAs for detecting ZEN in feed using this antibody. We suggest that the developed mAb and ELISA system can be effectively applied for the screening ZEN and its derivatives in animal feed and food.






 XML Download
XML Download