

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Transgenic plants have been used for the production and delivery of edible oral subunit vaccines [3,10]. Plant-based vaccines possess some advantages over other types of vaccine biotechnology such as increased safety, anticipated low cost of mass vaccination programs, and wider use of vaccines for veterinary medicine [9,10]. Several plant-derived vaccine candidates have been developed, and their immunogenicity and ability to protect against microbial infection were evaluated [3]. Corn, a major forage crop, is a particularly attractive expression vehicle due to its extensively studied genetics and amenability to established transformation procedures [7]. In addition, grain seeds provide excellent systems for oral delivery of subunit vaccines because of their low water content and long shelf-life [7].
Actinobacillus (A.) pleuropneumoniae is the etiological agent of severe hemorrhagic, necrotizing pneumonia often associated with fibrinous pleuritis in pigs, and is responsible for significant economic losses worldwide [2]. Among several virulence factors such as exotoxins, lipopolysaccharides, capsular polysaccharide, membrane proteins, and adhesion factors, the Apx exotoxin is believed to be correlated with A. pleuropneumoniae virulence [2]. Apx toxins are highly immunogenic and induce a substantial production of antibodies during A. pleuropneumoniae infection [4]. ApxII is expressed by all but serotype 10, while ApxI and ApxIII are expressed by different serotypes in various combinations [2]. In addition, serovars producing ApxI and ApxII are the most virulent. Therefore, ApxII could effectively serve as an antigen for vaccines against A. pleuropneumoniae. ApxIIA, which is expressed in Saccharomyces cerevisiae and transgenic tobacco, has previously been reported to be capable of inducing protective immune responses against A. pleuropneumoniae in mice [6,8,12]. The neutralizing epitope of ApxIIA (ApxIIA#5) from A. pleuropneumoniae serotype 2 was isolated in Korea and found to induce a protective immune response against A. pleuropneumoniae. Smaller fragments of this epitope are easily expressed in large quantities using a heterologous expression system [11].
Cholera toxin B (CTB) binds to GM1-ganglioside at the surface of mammalian intestinal epithelial cells, including the M cells of gut-associated lymphoid tissue [1]. The CTB subunit protein has also been found to induce mucosal immunity as an effective carrier molecule [1]. Therefore, it was chosen as a fusion protein for the present study.
The generation of transgenic corn expressing CTB-ApxIIA#5 fusion protein (0.93 kb) and full-size (2.8 kb) ApxIIA has been previously described [5]. One gram of the corns expressing the CTB-ApxIIA#5 fusion protein or full-size ApxIIA was ground in liquid nitrogen using a mortar and pestle. Soluble proteins were extracted at a solid-to-liquid ratio of 1 : 10 with phosphate buffered saline (PBS). The extracted protein was subcutaneously injected into mice to evaluate the immunogenicity of the transgenic corn-derived ApxIIA and CTB-ApxIIA#5 fusion proteins. Four-week-old ICR female mice (Orient Bio, Korea) were used through this study following policy and regulation for the care and use of laboratory animal (Laboratory Animal Center, Seoul National University, Korea). The mice were divided into three groups: a non-vaccinated control group and groups that received injections of the CTB-ApxIIA#5 fusion protein or full-size ApxIIA. Four mice in each group were boosted subcutaneously with 200 µL of soluble antigen extracts mixed with complete Freund's adjuvant (Sigma, USA), and were injected twice with the same antigen mixed with incomplete Freund's adjuvant (Sigma, USA).
Blood samples and spleens were collected 2 weeks after the final immunization. A spleen cell suspension was prepared by gently pressing and straining the spleen tissue through a 100-µm pore nylon cell strainer (100 µm; BD Falcon, USA) with a plunger. A 5 × 106 cells/mL suspension was cultured for 5 days in RPMI-1640 medium (Gibco-BRL, USA) supplemented with 10% fetal bovine serum (Gibco-BRL, USA) at 37℃ in 5% CO2. Spleen cells of each group were stimulated with the recombinant ApxIIA (2 µg/mL), and Concanavalin A (ConA, 2 µg/mL) to evaluate production of interferon (IFN)-γ, interleukin (IL)-4, and nitric oxide (NO).
After stimulation, the secreted concentration of IFN-γ and IL-4 was measured using the enzyme-linked immunosorbent assay (ELISA) kit (eBioscience, USA) according to the manufacturer's instructions. The levels of NO derived from the culture supernatant were determined by the Griess method [13]. ApxIIA-specific IgG and IgA-memory B cell enzyme-linked immunospot (ELISPOT) assays were performed using mouse IgG and IgA ELISPOT kits (Mabtech, USA) according to the manufacture's instruction.
The level of antigen-specific antibody in serum was determined with an ELISA. For this, 96-well microplates were coated with 1 µg/mL of the recombinant ApxIIA antigen and incubated overnight at 4℃. The plates were then blocked with 1% bovine serum albumin (Sigma, USA) in a solution of PBS containing 0.1% Triton X-100 (PBST) for 2 h at 37℃. After washing with PBST, 1 : 20 diluted mice sera were added to each well and incubated for 1 h at 37℃. Goat anti-mouse IgG (H + L)-HRP conjugate (Bio-Rad, USA) or anti-mouse IgA (α-chain specific)-HRP conjugate (Bethyl, USA) was used at 1 : 2,000 as a secondary antibody, and 2,2'-azin-bis (3-ethylbenzthiazoline-6-sulfonic acid) substrate (Bio-Rad, USA) was added to develop the color. Optical density was measured at 405 nm using an Emax Precision microplate reader (Molecular Devices, USA). Statistical significance was analyzed with Student's t-test using SPSS (ver 17.0; SPSS, USA). Differences were considered significant if a p-value < 0.05 was obtained.
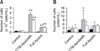
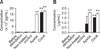
The level of ApxIIA-specific IgG antibodies significantly increased in serum from the mice boosted with the CTB-ApxIIA#5 fusion protein and full-size ApxIIA from the transgenic corn compared to the control mice at the time of the final booster shots (p < 0.01). However, there were no significant differences in the anti-ApxIIA IgA antibody responses between the injected groups and the control group (Fig. 1). The production of ApxIIA-specific IgG memory B cells was observed in spleens from the mice boosted with the CTB-ApxIIA#5 fusion protein and full-size-ApxIIA by ELISpot (p < 0.01) from the transgenic corn (Fig. 2). In addition, ApxIIA-specific IgA memory B cells were produced in the mice injected with the CTB-ApxIIA#5 fusion protein (p < 0.05; Fig. 2).
The secretion of IFN-γ, IL-4, and NO from spleen cells was measured as an indicator of immune response induction. After in vitro stimulation, spleen cells from the group injected with full-size ApxIIA produced high levels of IFN-γ, a T helper type 1 cytokine (p < 0.05; Fig. 3), while the production of IL-4, a T helper type 2 (Th2) cytokine, was not detected in any group (data not shown). The transgenic corn-derived ApxIIA, particularly the CTB-ApxIIA#5 fusion protein, was found to induce the production of antigen-specific antibodies (Figs. 1 and 2), although IL-4 related to a Th2 immune response was not detected. In addition, high NO production was detected in the mice treated with the CTB-ApxIIA#5 fusion protein and full-size ApxIIA after in vitro stimulation (p < 0.01; Fig. 3).
Based on the induced antigen-specific immune responses, we concluded that the transgenic corn-derived CTB-ApxIIA#5 fusion protein and full-size ApxIIA are capable of effectively inducing Apx-specific immune responses in mice. In particular, the group injected with the CTB-ApxIIA#5 fusion protein showed higher ApxIIA-specific antibody titers than the group injected with the full-size ApxIIA. Fusing the ApxIIA#5 antigenic fragment with the CTB protein could be thought to improve the efficiency of antigen delivery to the host immune system based on these results. Moreover, the small size of the ApxIIA epitope contributes to increased expression levels in a heterologous expression system [11]. Consequently, the transgenic corn expressing the CTB-ApxIIA#5 fusion protein and full-size ApxIIA is a promising candidate for an effective vaccine against A. pleuropneumoniae. We are currently preparing for further experiments to develop a transgenic corn-derived oral vaccine against A. pleuropneumoniae based on the results of the present study.

 XML Download
XML Download