

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Ticks are of great veterinary importance compared to other ectoparasites. These parasites consume large quantities of host blood during their lengthy attachment period (7~14 days), which may be extended depending on the tick species and unique host association. The control of tick infestation has centered on the use of acaricides and on animal breeds that have some degree of resistance against ticks. Continuous use of acaricides leads to the appearance of acaricide-resistant ticks [18]. There is also increasing evidence that the strategies that rely heavily on acaricides are not cost-effective [20]. These factors, in addition to the concerns about chemical residues in livestock products and the undesirable effects of chemicals in the environment, have led to the search for alternative methods of tick control. Host immunization is one of the most promising options [7,21,22].
The potential of an anti-tick vaccine for reducing tick populations [7] and limiting the transmission of certain diseases has been well-documented [7,9]. Immunization of animals with tick-protective antigens is promising and attractive method as it is not associated with any health risks, and is environmentally safe and cost-effective [28]. Identification of new antigens from various tick species is expected to contribute the development of more effective vaccines or polyvalent vaccines that are effective against numerous tick species [29].
Hyalomma (H.) dromedarii can behave as a three- or two-host species, with the two-host life cycle seemingly the most common. Camels are the main hosts of the adults, which also parasitize other domestic animals. Nymphs and larvae can parasitize the same hosts as adults, especially camels, but rodents, hedgehogs, and birds can also serve as hosts [3]. H. dromedarii is widely distributed throughout North Africa, the northern regions of West, Central, and East Africa; Arabia, Asia Minor, the Middle East, and Central and South Asia [4]. The aim of the present study was to isolate and characterize immunogenic glycoproteins (GLPs) from the adult and larval H. dromedarii. We then tested the protective effects of immunization with the isolated GLPs against experimental infestation challenges of this tick species.
Materials and Methods
Tick specimens
Engorged females and nymphs of the camel tick H. dromedarii were originally collected from the ground of camel pens in Burkash village, Giza governorate, Egypt. Identification of females was confirmed in the laboratory according to the keys of Hoogstraal [16] and Estrada-Peña et al. [10]. The females were incubated at a constant temperature of 27 ± 2℃ with a relative humidity of 75 ± 5% in permanent darkness to obtain eggs and larvae as previously described [19]. Engorged nymphs were also incubated under the same conditions until they molted to the next instar (unfed adults). Unfed adults were divided into two groups: a group used as a source of antigenic material and another group used for challenge infestations. Larvae were only used as a source of antigenic material.
Preparation of whole adult and larval antigens
Whole adult and larval antigens of H. dromedarii were prepared according to the method of Ghosh and Khan [12]. In brief, laboratory-reared, 5- to 6-day-old unfed ticks or larvae were homogenized in cold 0.15 M phosphate-buffered saline, pH 7.2, containing 1 mM disodium EDTA and a protease inhibitor cocktail (Sigma, USA). The homogenate was then filtered, sonicated in ice for 3~4 cycles for 20~25 sec each at 16-micron amplitude, and centrifuged at 15,000 × g for 60 min at 4℃. The supernatants were designated as whole adult or larval antigen according to the source. Total protein concentrations of the supernatants were estimated according to the Bradford method [5].
Concanavalin A (Con-A) affinity chromatography
Whole adult and larval antigens of H. dromedarii were equilibrated with 20 mM phosphate buffer (PB) containing 0.5 M NaCl, pH 7.4, and loaded onto a Con-A sepharose column (1.6 × 4 cm; GE Healthcare, Sweden). The unbound proteins were washed with equilibration buffer at a flow rate of 30 mL/h. The bound proteins were eluted with 0.2 M methyl α-D glucopyrinoside at a flow rate of 30 mL/h. The eluted proteins were dialyzed against 20 mM PB containing 0.5 M NaCl, pH 7.4, and designated as whole adult GLPs (AGLPs) or larval GLPs (LGLPs). Total protein content of the GLPs was estimated by the Bradford method [5].
SDS-PAGE
Electrophoretic analysis was performed using the Mini-Protean II Dual-Slab Cell (BioRad, USA). Preparation of the gels and samples, and electrophoresis were performed according to the method described by Laemmli [17].
Immunization schedule
Twelve Chinchilla bastard rabbits (male, body weight 3 kg; NRC Animal Facility, Egypt) were divided into three groups: four rabbits were immunized with AGLPs, four rabbits were immunized with LGLPs, and four rabbits were immunized with saline (as a negative control). All rabbits were immunized by intramuscular injection of 20 µg AGLPs or LGLPs in 0.5 mL saline mixed with an equal volume of Freund's complete adjuvant (Sigma, USA) on day 0. Rabbits were boosted with another intramuscular injection of 20 µg of the same antigen mixed with Freund's incomplete adjuvant (Sigma, USA) on day 14 and 28. Ten days after boosting, the rabbits were bled from the marginal ear vein. The antisera were collected and pooled for immunoblotting assays.
Immunoblotting
Immunoblot analysis was performed using a NovaBlot semi-dry blotter (LKB Produkter, Sweden). Preparation of buffers, samples, and the transfer procedure was carried out according to the method described by Towbin et al. [26] with slight modifications. Rabbit anti-GLP antisera was used at dilution of 1:1,000 in 0.01 M Tris buffered saline, pH 7.5, containing 0.5% bovine serum albumin. Primary antibody binding was visualized with an anti-rabbit IgG peroxidase conjugate (Sigma, USA) at 1:3,000 dilution in the same buffer for 1 h at 37℃, and 4-chloro-1-naphthol as a substrate.
Challenge infestation with adult H. dromedarii ticks
Immunized and control rabbits were exposed to adult H. dromedarii ticks. The ticks were placed inside a feeding capsule consisting of a plastic tube (2.5 cm of diameter and 3 cm of height) glued to the shaved backs of the rabbits (two capsules per animal). Wooden collars were placed on the rabbits to prevent grooming as previously described [25]. Twenty adult ticks (10 in each capsule; 5 females and 5 males) were placed on each rabbit. Ticks were monitored daily to observe their feeding habits. Biological parameters such as feeding period, body weight of the engorged females, egg mass, and egg hatching were recorded. The reproductive index was calculated as described by Singh and Ghosh [24].
Results
Purification of whole AGLPs and LGLPs
AGLPs and LGLPs from H. dromedarii ticks were purified from 60 mg and 130 mg of adult and larval proteins, respectively, using affinity chromatography on a Con-A sepharose column. The total amount of purified whole AGLPs recovered was 1.7 mg while the total amount of purified LGLPs was 4.7 mg (Figs. 1 and 2).
Characterization of whole AGLPs and LGLPs
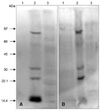
Electrophoretic analysis of whole adult and larval H. dromedarii proteins showed several proteins of high, medium, and low molecular weights. The electrophoretic patterns of whole AGLPs and LGLPs included three major protein bands around 97, 66 and 40 kDa in size (Fig. 3).
Immunoblot analysis of the purified LGLPs
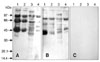
Chinchilla bastard rabbits were immunized three times with either the purified AGLPs or LGLPs. Sera were collected before injection and 2 weeks after each injection. The rabbit anti-AGLPs and anti-LGLPs were isolated and their reactivity against the whole H. dromedarii adult tick, H. dromedarii larval antigens, AGLPs and LGLPs proteins was determined by immunoblotting (Fig. 4). Three major protein bands with molecular weights of about 97, 66, and 40 kDa corresponding to the purified GLPs were observed (Figs. 4A and B). These results showed that the antisera raised against AGLPs and LGLPs contain specific antibodies against these GLPs.
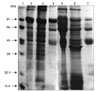
The rabbit anti-H. dromedarii AGLPs and anti-H. dromedarii LGLPs detected different proteins from the adult and larval H. dromedarii ticks. Proteins and purified GLPs of adult and larval H. dromedarii ticks showed no reactivity with normal rabbit serum (Fig. 4). Rabbit anti-H. dromedarii AGLP and anti-H. dromedarii LGLP detected bands of Rhipicephalus (R.) sanguineus proteins with different molecular weights (Figs. 5A and B, lane 2). This observation demonstrates the cross-reactivity between H. dromedarii and this species. Both antisera failed to detect specific bands of proteins extracted from the soft tick O. moubata (Figs. 5A and B, Lane 3). These findings indicate that GLPs from both whole adult H. dromedarii ticks and larvae might be useful in the vaccination against R. sanguineus infestations.
Feeding and reproductive performance of adult females
Table 1 shows the immunological protection from immunization with AGLPs and LGLPs isolated from unfed adult and larval H. dromedarii and the effect on feeding and reproduction of adult females. Both antigens AGLPs and LGLPs did not record any rejection as that in control. Females fed on immunized rabbits showed insignificant increases in their feeding period compared to ones fed on control animals. Immunization with both AGLPs and LGLPs caused insignificant increases in the engorged weight of females. Egg masses were insignificantly reduced by immunization with AGLP and insignificantly increased with LGLP immunization. Reproductive index associated with AGLP (p < 0.01) and LGLP (p < 0.05) immunization was significantly lower compared to the control. Egg hatching rates were significantly reduced by immunization with both antigens. Generally, the two tested antigens had modest effects on reproductive index and strongly affected egg hatching.
Discussion
Ticks are distributed throughout the world and impact human and animal health as well as food animal production. Control of ticks has been primarily achieved by the application of acaricides, which has resulted in the selection of resistant ticks and environmental pollution. Vaccines have been shown to be a feasible method of tick control that offers a cost-effective and environmentally-friendly alternative to chemical control. However, identification of tick-protective antigens remains the limiting step in vaccine development. Tick antigens naturally exposed to the host during tick feeding and ones that are concealed have both shown promise as candidate vaccine antigens. Development of vaccines against multiple tick species may be possible using highly conserved tick-protective antigens or antigens that show immune cross-reactivity among different tick species.
Vaccines made from a combination of key protective antigens may greatly enhance vaccine efficacy. Previous studies have suggested the use of vaccine strategies that prevent both tick infestation and pathogen transmission [9]. The idea of developing immune-based prophylactic measures against infestation with multiple breeds of ticks on crossbred animals is based on the concept that ticks feeding on immunized hosts may ingest antibodies specific for a target antigen(s) within the tick, thereby exerting deleterious effects on feeding and reproduction [2,21].
Infestation with multiple tick species may prevent the efficacy of tick vaccines with a narrow spectrum that are produced with antigens from a single tick species. Recombinant BM86 included in commercial vaccine formulations including Tick-GARD and Gavac confers partial protection against ticks of the phylogenetically-related Hyalomma and Rhipicephalus generas. However, immunization with BM86 fails to protect against the more phylogenetically distant Amblyomma spp.. Therefore, identification of broad-range vaccine candidates against infestations with ticks of phylogenetically-distant species is needed [15].
Immunoaffinity chromatography was previously used to purify salivary gland antigens from Amblyomma americanum [6], larval antigens [12-14], gut larval antigens [8], and nymphal antigens [23] of H. anatolicum and Boophilus (B.) microplus [11]. The most important factors that influence immunoprotective effects are the antigenic dose and combination with specific adjuvants [24]. Willadsen et al. [27] examined the effects of two doses (9.2 and 17.0 µg) of 89 kDa GLPs per animal in two separate experiments, and found that this formulation provided protection against B. microplus challenge infestations. In another study, Australian scientists used 500 µg of affinity-purified larval antigens per animal [30] and vaccinated cattle were protected (78%) against challenge with B. microplus tick larvae.
For H. anatolicum, 2 mg of larval 39 kDa protein [12,13], 1.6 mg of gut larval antigens [8], and 1.6 mg of nymphal 39 kDa protein [23] were found to be protective against homologous challenge. In the present investigation, a biochemical approach was used to purify the major GLPs of the H. dromedarii tick. Three major GLPs with molecular weights of approximately 97, 66, and 40 kDa were purified using a single-step purification method with a Con-A sepharose column. These GLPs reacted with the rabbit antisera against both adult and larval H. dromedarii antigens, which demonstrates that these GLPs have specific antibodies in these antisera. Our results suggest that these GLPs may be suitable immunogens for the vaccination of camels against tick infestation. The immunogenicity of these proteins could be a result of their role as membrane proteins. Reaction of these GLPs with sera from hyperimmune rabbits would confirm the fact that unfed larvae are a good source of biological material for the isolation of protective antigens, and would be consistent with data presented by Ghosh and Khan [12] and Ghosh et al. [13], who confirmed that 39 kDa immunoaffinity chromatographgy purified protein from unfed larvae of H. anatolicum anatolicum provided a relatively easily available source of antigen for immunization of cattle against both larvae and nymphs of this species.
More interestingly, rabbit antisera against these GLPs reacted with proteins from other tick species in the immunoblot assay. This demonstrated the immunogenicity of GLPs isolated from either adult or larvae. Additionally, these antisera failed to detect bands in the protein extracts of R. annulatus (formerly B. annulatus) while proteins from R. sanguineus were detected although both are of these species are hard ticks.
The efficacy of immunization with the purified GLPs was confirmed by the challenge experiment which demonstrated 63~67% reduction in egg hatchability. Unlike Das et al. [8], Abdel-Shafy et al. [1] noted significant changes in the percent of rejection, feeding period, and egg mass when they used purified or fractionated antigens from larvae of H. anatolicum and guts of H. dromedarii. Our results did not show any significant changes in these parameters. Our findings might be attributed to the low inoculation dose (20 µg) of either the AGLPs or LGLPs used for rabbit vaccination. This conclusion was made based on the results of the infestation challenge test that showed immunization had significant effects on only egg hatchability whereas all other biological parameters were similar to those of the control group. Das et al. [8] used 300 and 650 µg of affinity purified gut antigens of H. anatolicum anatolicum to inoculate calves and Abdel-Shafy et al. [1] inoculated experimental rabbits with 300 µg of gel filtration gut antigens of H. dromedarii, recording significant effects with most of the previously mentioned biological parameters. Therefore, it is believed that these significant effects might be attributed to the use of high doses of tick antigens. The insignificant effects on engorgement weight of females observed in this study agree with those reported by Das et al. [8] and Abdel-Shafy et al. [1]. The reduction in egg hatchability we observed also agrees with that reported by Abdel-Shafy et al. [1] in a study that used fractionated gut antigens of H. dromedarii to protect against tick infestation. Thus, we can hypothesize that immunization with the purified GLPs has extended effects on subsequent generations of ticks or egg viability.
In conclusion, three major GLPs were purified from adult and larval H. dromedarii camel ticks and were evaluated as vaccines against camel tick infestation using a rabbit model. The reproductive index was modestly affected by immunization with these GLPs while egg hatchability was significantly reduced. Our study showed that immunization with the purified GLPs provides protection against H. dromedarii infestation, but further studies are needed to examine the effects against other tick species.



 XML Download
XML Download