

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Ionizing radiation is a very useful tool in a wide variety of fields including medicine, agriculture, industry, research, and military operations. However, it has various effects on both the developing and adult brain, even though the adult brain displays less severe morphological changes to irradiation than other organs. A relatively low-dose of whole body exposure in experimental animals can produce transient damage to immature progenitor cells in specific tissues, including the brain [7]. Especially, under very low doses of radiation, the developing brain is one of the most radiosensitive organs [6,10].
In adults, although irradiation is one of the most effective therapies for treatment of head and neck tumors, this therapeutic application is limited due to potential damage of normal brain tissue [9]. Ionizing radiation induces apoptosis of neural immature progenitor cells in the subventricular zone of the lateral ventricles and in the subgranular zone of the hippocampal dentate gyrus in the adult brain [3,7]. The phenomenon in the adult mouse hippocampus may lead to hippocampal-dependent learning and memory impairment [7]. Even very low doses of irradiation to the brain of an infant before age 18 months can affect the patient's cognitive abilities in adulthood [5]. However, mechanistic studies of radiation-induced effects on cognitive function related to the hippocampus are very difficult because they rely on complicated in vivo models [2]. A previous in vitro study confirmed that the radiosensitivity of immature hippocampal cells is significantly higher than that of the mature cells, indicating that the susceptibility of such hippocampal cells depends on their differentiation [12]. Therefore, further study needs to address the specific mechanisms associated with radiation effects on immature hippocampal neurons by using in vitro approaches.
This study examined the cytotoxicity of acute gamma (γ)-irradiation in immature hippocampal neurons to clarify the precise mechanism(s) in the detrimental effect of ionizing radiation on immature hippocampal neurons in an in vitro system.
Materials and Methods
Primary hippocampal neuron culture
Pregnant Sprague-Dawley rats were obtained from a specific pathogen-free colony at Oriental Inc. (Korea). The method used to primary hippocampal neuron culture has been previously described in detail [12]. Briefly, hippocampi were dissected from postnatal day 0 rats. After dissection, tissues were chopped and digested with 10 units/mL papain (Worthington, USA) and 100 units/mL DNase I (Roche, Switzerland) in dissociation buffer at 37℃ for 30 min. The digestion was triturated with Neurobasal A medium (Invitrogen, USA). The cells were seeded at a density of 4 × 106 cells/well on poly-D-lysine hydrobromide (150 µg/mL; Sigma-Aldrich, USA) coated 4-well plates. Two hours after plating, Neurobasal A was replaced with growth medium including Neurobasal A, 1 × B27 supplement (Invitrogen, USA), 100 units/mL penicillin, 0.1 mg/mL streptomycin, and 0.5 mM glutamine.
Double-immunofluorescent staining
Hippocampal cultured neurons [0.5 day in vitro (DIV)] on a cover slip were fixed with 4% paraformaldehyde (Sigma-Aldrich, USA) in phosphate-buffered saline (PBS, pH 7.4) for 20 min at room temperature. The cells were blocked with 2% normal goat serum and 1% bovine serum albumin, allowed to react with rabbit monoclonal anti-Ki-67 (DRM004, 1 : 200; Acris Antibodies GmbH, Germany) and rabbit polyclonal anti-doublecortin (DCX) antibody (1 : 400; Cell Signaling Technology, USA) overnight at 4℃, and incubated with tetramethylrhodamine isothiocyanate-conjugated goat anti-rabbit IgG (1 : 50; Sigma-Aldrich, USA) for 1 h at room temperature. The cells were incubated with either mouse monoclonal anti-nestin (1 : 200; Chemicon International, USA) or mouse monoclonal anti-glial fibrillary acidic protein (GFAP) antibody (1 : 500; Sigma-Aldrich, USA) for 1 h at room temperature, and then incubated with fluorescein isothiocyanate-conjugated goat anti-mouse IgG (1 : 50; Sigma-Aldrich, USA) for 1 h at room temperature. For counterstaining, 1 mg/mL 4',6-diamidino-2-phenylindole 2HCl (DAPI; Thermo Fisher Scientific, USA) in PBS was added and incubated at room temperature for 5 min. The immunofluorescence-stained specimens were examined using a ProgRes CFscan fluorescence digital camera (Jenoptik, Germany).
Irradiation and treatment
Cultured neurons at 0.5 DIV were irradiated with 0, 0.5, 2, or 4 Gy of 137Cs-generated γ-rays using a Gamma-cell Elan 3000 (Nordion International, Canada). The mean dose rate of the γ-rays was 3.1 Gy/min. Control samples were sham-irradiated with 0 Gy. The cells were assayed at 24 h post-irradiation. To evaluate the radioprotective effects of anti-oxidative substances, including amifostine (0~100 µM; a representative agent of radioprotactants; Sigma-Aldrich, USA) and epigallocatechin gallate (EGCG, 0~10 µM; the main ingredient of green tea polyphenols; Sigma-Aldrich, USA) on immature hippocampal neurons, both substances were pre-treated at 30 min before irradiation.
Cytotoxicity and reactive oxygen species (ROS) evaluation
Cytotoxicity was evaluated using a lactate dehydrogenase (LDH) release assay. A commercially available LDH-cytotoxicity assay kit from Biovision (USA) was used as recommended by the manufacturer. The optical density values were quantified by an Emax microplate reader (Molecular Devices, USA) for absorbance at a wavelength of 490 nm. To monitor intracellular accumulation of ROS, the fluorescent probe 2',7'-dichlorodihydrofluorescein diacetate (H2DCF-DA; Invitrogen, USA) was used. Intracellular ROS production was quantified by a fluorescent microplate reader with excitation at 485 nm and emission at 535 nm.
Western blotting
Medium was removed and sodium dodecyl sulfate (SDS) sample buffer (4×) was added to each culture. The cells were scraped and sonicated for 10 sec. The samples were heated to 100℃ for 10 min. The samples were then separated by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE), and the resolved proteins were transferred to a nitrocellulose membrane and blocked with 1% normal goat serum and 0.5% bovine serum albumin in PBS-T (PBS, 0.1% Tween 20) for 30 min at room temperature. The membranes were then incubated with a 1 : 1,000 dilution of either rabbit anti-cleaved (active) caspase-3 antibody (Cell Signaling Technology, USA) or rabbit anti-poly (ADP-ribose) polymerase (PARP) antibody (Cell Signaling Technology, USA) in PBS-T overnight at 4℃. After extensive washing and incubation with a 1 : 10,000 dilution of horseradish peroxidase-conjugated goat anti-rabbit IgG (Vector Laboratories, USA) for 1.5 h at room temperature, the signals were visualized using chemiluminescence methods (SuperSignal West Pico; Thermo Fisher Scientific, USA). For normalization purposes, the membranes were reprobed with a 1 : 20,000 dilution of β-actin antibody (Sigma-Aldrich, USA). Several exposure times were used to obtain the signals in the linear range. The bands were quantified using Scion Image Beta 4.0.2 for Windows XP software (Scion, USA).
Results
Cell phenotypes of 0.5 DIV hippocampal culture
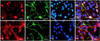
To confirm the cell phenotypes of 0.5 DIV hippocampal cultured cells, double-immunofluorescent staining was performed. We first detected Ki-67, nestin, DCX and GFAP, which are immunohistochemical markers for proliferating cells, neural stem cells, immature progenitor neurons and astrocytes, respectively, in 0.5 DIV hippocampal cells. The double-immunofluorescent results showed that Ki-67-positive cells were co-localized to nestin immunoreactivity (approximately 15% of total cells; Figs. 1A~D), but DCX-positive cells (approximately 80% of total cells) were rarely matched with GFAP immunoreactivity (< 5% of total cells; Figs. 1E~H) in 0.5 DIV hippocampal cells.
Cytotoxic effect of γ-rays on immature hippocampal neurons
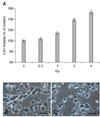
The cytotoxicity of γ-irradiation in immature hippocampal cultured neurons was estimated 24 h after irradiation by 0~4 Gy by the LDH release assay (Fig. 2). The cytotoxicity increased in a dose-dependent pattern within the range of irradiation applied.
Caspase-dependent cytotoxicity of γ-ray in immature hippocampal neurons
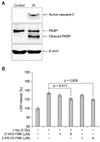
To determine whether radiation-induced cell death in immature hippocampal neurons was associated with the caspase-dependent pathway, caspase-3 activation and caspase-specific PARP cleavage were assessed by Western blot and the effects of caspase inhibitors on radiation-induced cytotoxicity were examined by LDH release assay (Fig. 3). Active caspase-3 and cleaved PARP markedly increased in immature hippocampal neurons at 24 h after 2 Gy γ-irradiation (Fig. 3A). However, both the caspase-family inhibitor Z-VAD-FMK and the caspase-3 specific inhibitor Z-DEVD-FMK significantly blocked LDH release from γ-irradiated immature hippocampal neurons (Fig. 3B).
Radioprotective effect of anti-oxidative substances in immature hippocampal neurons
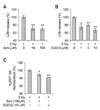
We further examined whether anti-oxidative substances, including amifostine (well-known as a representative radioprotective agent [8]) and EGCG could inhibit the cytotoxicity of γ-rays in immature hippocampal neurons. Pre-treatment of amifostine and EGCG significantly blocked increase of LDH release (Figs. 4A and B) and intracellular ROS levels (Fig. 4C) caused by 2 Gy-irradiation.
Discussion
In this study, we demonstrate the cytotoxicity of γ-rays in immature hippocampal neurons via the caspase-dependent pathway. Further, we suggest that the caspase-dependent cytotoxicity of γ-rays in immature hippocampal cultured neurons may be caused by oxidative stress.
An irradiation study using developing rat brains suggested that the radiosensitive population consists primarily of cells nearing cell division or cells that have recently completed mitosis and are beginning to differentiate [1]. Viability of immature hippocampal cells, but not mature cells, can significantly decline upon exposure to γ-rays, indicating that neuronal resistance to γ-irradiation is associated with neuronal differentiation [11,12]. Furthermore, cultured hippocampal cells form functional synapses and a network after 3~5 DIV [13]. Therefore, we chose to apply γ-irradiation to 0.5 DIV-cultured hippocampal neurons, which are comparable to neural immature progenitor neurons before cell connection and synapse formation.
In addition, this study detected immunohistochemical markers for proliferating cells, neural stem cells, immature progenitor neurons, and astrocytes in 0.5 DIV hippocampal cells. This suggests that rat immature hippocampal cells in this in vitro system, which have > 95% immature progenitor neural cells, can be very useful for screening radioprotectants and studying various neuronal perturbation factors caused by radiation exposure.
Exposure of mammalian brains to ionizing radiation induces apoptosis of neural immature progenitor cells in vivo [3]. DNA-fragmentation and apoptotic appearance in immature hippocampal neuron are increased by γ-irradiation in a dose-dependent manner [12]. This study also showed that γ-ray-induced cytotoxicity in immature hippocampal neurons was increased in a dose-dependent pattern within the range of irradiation applied. Additionally, active caspase-3 and cleaved PARP were markedly increased in immature hippocampal neurons with 2 Gy γ-irradiation. Furthermore, both the caspase-family inhibitor Z-VAD-FMK and the caspase-3 specific inhibitor Z-DEVD-FMK significantly blocked radiation-induced cytotoxicity in immature hippocampal neurons. This suggests that γ-irradiation-induced cell death in immature hippocampal neurons depends on a pro-apoptotic caspase pathway, especially the caspase-3 related pathway.
Exposure of ionizing radiation to the central nervous system induces neuronal apoptosis via oxidative stress [4]. Appropriately, an experiment examined whether anti-oxidative substances could inhibit the γ-ray-induced cytotoxicity in immature hippocampal neurons. In this study, pre-treatment of amifostine and EGCG significantly blocked γ-ray-induced LDH release and intracellular ROS levels. These results support the suggestion that oxidative stress is associated with the cytotoxicity of γ-rays in immature hippocampal cultured neurons. Additionally, this experiment confirmed a beneficial in vitro system for screening radioprotective compounds.
Collectively, this in vitro study has confirmed that the caspase-dependent cytotoxicity of γ-rays in immature hippocampal cultured neurons may be caused by oxidative stress. In addition, this study provides a useful tool for the analysis of critical mechanistic parameters associated with radiation response of the brain and to better understand the molecular changes in radiation-induced cell death of immature neural cells.
 XML Download
XML Download