

 PDF
PDF ePub
ePub Citation
Citation Print
Print


β-glucans are heterogeneous glucose polymers that form a structural extracellular matrix in pathogenic microbes [1]. These compounds are recognized by several receptors on the surface of immune cells [4]. Recent studies demonstrated that a novel β-glucan purified from Paenibacillus (P.) polymyxa JB115 [8] induces macrophage activation [2].
Dendritic cells (DCs) are the most potent antigen-presenting cells in the immune system [12]. They are specialized for capturing and processing antigens, then presenting antigenic peptides to naïve T lymphocytes to initiate antigen-specific immune responses [13]. However, the biological effects of β-glucan purified from P. polymyxa JB115 on DCs remain unclear. In the present study, we evaluated the various biological effects of β-glucan on DCs.
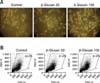
To investigate the effects of β-glucan purified from P. polymyxa JB115 on DCs, we prepared immature DCs from bone marrow as previously described [10], and treated these cells with 1~100 µg/mL β-glucan. Two days after β-glucan treatment, clusters of DCs were observed using an inverted microscope connected to a digital camera (Fig. 1A). In addition, cell size was analyzed using flow cytometry. It was found that β-glucan enhanced the percentage of normal-sized DCs in the forward scatter/side scatter dot plot (Fig. 1B). A cell viability assay using a trypan blue solution demonstrated that 100 µg/mL β-glucan significantly increased the viability of DCs (Fig. 2). Thus, β-glucan may enhance the survival of DCs and also helps maintain normal cell size.
The production of tumor necrosis factor (TNF)-α and interleukin (IL)-12 by DCs treated with β-glucan was determined by an ELISA. IL-12 and TNF-α are representative cytokines involved in cell-mediated and innate immunity, respectively, along with DC survival [3,5]. β-glucan (100 µg/mL) significantly increased the production of both cytokines (Figs. 3A and B). In addition, β-glucan enhanced the release of nitric oxide from DCs (Fig. 3C). Nitric oxide plays a critical role in eliminating intracellular pathogens in macrophages.
To determine whether β-glucan induces the maturation of DCs, expression levels of immune-related DC surface markers were measured by flow cytometric analysis [9]. Major histocompatibility complex (MHC) class I and II are major antigen-presenting molecules on DCs while CD54 and CD86 are adhesion and co-stimulatory molecules, respectively [11]. β-glucan consistently enhanced the expression all of the surface markers investigated in the present study (Fig. 4A). Because MHC class II and CD86 molecules have been identified as maturation markers of bone marrow-derived DCs [6,12], β-glucan may induce the maturation of DCs. Decreased antigen uptake is a major characteristic of mature DCs. Antigen uptake analysis using dextran-fluorescein isothiocyanate revealed that β-glucan profoundly decreased the antigen uptake capability of DCs compared to immature DCs (Fig. 4B). Zymosan was used as a positive control in this assay.
To confirm the antigen-presenting capability of DCs treated with β-glucan, we prepared allogeneic splenocytes from BALB/c mice [7]. The cells were then stained with carboxyfluorescein succinimidyl ester (CFSE) and co-cultured with immature or β-glucan-treated DCs. Highly proliferative cells had a relatively low CFSE intensity detected as FL1 by flow cytometric analysis because of their high cell division rates. Splenocytes cocultured with DCs treated with β-glucan (100 µg/mL) showed the lowest CFSE intensity, indicating that these cells had the highest proliferative capacity (Fig. 5). These observations showed that β-glucan enhances the antigen-presenting capability of DCs. Taken together, our results demonstrated that β-glucan from P. polymyxa JB115 induces the maturation of DCs, the most potent antigen-presenting cells in host immunity. This study may provide researchers with valuable information to increase the use of β-glucan.




 XML Download
XML Download