

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Avian metapneumovirus (aMPV) is a member of the subfamily Pneumovirinae in the family Paramyxoviridae [26]. aMPV, also known as turkey rhinotracheitis virus, causes a widespread disease of turkeys, chickens and some other avian species [7]. The disease is characterized by an upper respiratory tract infection in turkeys. aMPV is also considered to be a predisposing factor for triggering swollen head syndrome (SHS) in broiler and broilers breeders [11, 13], and egg production losses in layers [29]. In chickens, it is well-known that the complication of secondary bacterial infections with organisms like Escherichia coli [10] and the immunosuppressive effect of infectious bursal disease [24] are important in the development of typical SHS.
Since aMPV was first described in South Africa in 1978 [3], the virus has been reported in other parts of Europe and the Middle East, and is now considered to be a major disease threat in both turkeys and chickens in many parts of the world [1,2,5,23,27]. Several subtypes of aMPV have been recognized and differentiated by nucleotide sequence analysis based on the attachment (G) protein [14]. Subtype A was first isolated in South Africa and England, whereas subtype B virus was initially isolated in continental European countries including Hungary, Spain and Italy. Subtype C was isolated in the United States [27], while the presence of an additional subtype D was reported in France [2].
In Korea, SHS was first observed in 1992 [16] and antibodies to aMPV have been detected in 21 out of 30 broiler breeder flocks (70%) [21]. Although clinical diseases similar to SHS have continuously reported in chicken flocks by field veterinarians, there have been no reports isolating and identifying aMPV subtype A in Korea. Recently, aMPV subtype C was isolated from pheasants in Korea [20].
To investigate the prevalence of aMPV in chickens in Korea, we developed a multiplex real-time reverse transcription polymerase chain reaction (RRT-PCR) for rapid detection of aMPV subtypes A and B. Although various methods such as a RT-nested PCR assay have been used to detect aMPV [7], it is time-consuming and sometimes produces false positive results caused by DNA contamination during secondary PCR. Here, we examined oropharyngeal swab and nasal turbinate samples taken from aMPV monitoring chicken farms using multiplex RRT-PCR and tried to isolate aMPV by Vero cell passages. For further analysis of the molecular and pathogenic characteristics of subtype A aMPVs isolated in this study, we analyzed the phylogenetic trees and investigated the pathogenicity of a Korean aMPV isolate in chickens and turkeys.
Materials and Methods
Preparation of clinical samples
Clinical samples were received for diagnosis and surveillance for aMPV during 2004-2008. A total of 130 chicken flocks (26 broiler, 87 broiler breeder and 17 layer flocks) with or without clinical signs ranging from mild to severe were examined. The oropharynx of six birds was swabbed per flock and each of two samples was pooled for this study. Oropharyngeal swabs or nasal turbinates were resuspended in minimum essential medium with gentamicin (Kukjae Pharmaco, Korea), the suspension was clarified by centrifugation at 1,000 × g for 10 min, and the supernatant was used immediately for RNA extraction and aMPV isolation.
aMPV reference viruses and cells
aMPV subtype A (Nobilis TRT vaccine; Intervet, The Netherlands) and subtype B (Nemovac; Merial, USA) vaccine strains were used as reference for both multiplex RRT-PCR and RT-PCR. Vero cells were prepared for aMPV isolation.
RNA extraction, multiplex RRT-PCR and RT-PCR
RNA was extracted directly from oropharyngeal swab or nasal turbinate samples using an RNeasy kit (Qiagen, USA) according to the manufacturer's instructions. To detect aMPV in samples, multiplex RRT-PCR was developed with primers and probes designed for this study (Table 1). QuantiTect Probe RT-PCR kit (Qiagen, USA) was used for the multiplex RRT-PCR reaction. Each reaction consisted of 10 µL total RNA, 12.5 µL QuantiTect master mix, 0.25 µL QuantiTect RT mix, 0.375 µL forward and reverse primers (each 55 µM) and 0.375 µL probes (each 14 µM), for a final volume of 25 µL. The multiplex RRT-PCR was performed on a SmartCycler (Cepheid, USA) with the following conditions: one cycle at 50℃ for 30 min, 95℃ for 15 min followed by 45 cycles of 94℃ for 15 sec, 55℃ for 60 sec. For the sequencing, RT-PCR was performed with primers as described by Cavanagh et al. [4]. The RT-PCR products were amplified for 268 base pair (bp) or 361 bp fragments corresponding to subtypes A and B, respectively, of the aMPV G gene.
Isolation of aMPV
Oropharyngeal swab or nasal turbinate samples confirmed to be aMPV positive by RRT-PCR were directly inoculated and passaged in Vero cells until cytopathic effects (CPE) were evident. When the Vero cells showed CPE, the aMPV was identified by immunofluorescence assay with the monoclonal antibody TRT12 (provided by National Veterinary Research and Quarantine Service, Korea) and confirmed by transmission electron microscopy.
Sequence analysis
To analyze the G glycoprotein from aMPV-positive samples, RT-PCR products (268 bp and 361 bp) were purified with the QIAquick PCR purification kit (Qiagen, USA) and sequenced by automated DNA sequencing using an ABI 377 apparatus (Applied Biosystems, USA). Sequences were aligned with the aMPV vaccine strains (Nobilis TRT vac/A and Nemovac/B) and previously published sequences: CVL14/1 (accession number L34032) and 8544 from England (DQ666911), 1556 from France (L34030), LAH (AY640317), SHSBR662/03 (AY739717), SHSBR668/03 (AY739718), SHSBR669/03 (AY739719), TRTBR169 (AY734531), chicken/A/BR/119/95 (AY842242), chicken/A/BR/121/95 (AY842243), aMPV/ B/Brazil-07/ USP-06 (EU140747), aMPV/B/Brazil-06/USP-07 (EU140748) and aMPV/B/Brazil-07/ USP-08 (EU140749) from Brazil, 872S from Spain (L34034), 6574 from Hungary (L34033), 2119 from Italy (L34031), Isr/1708/02 from Israeli (AY728268), Colorado 2 from United States (AY590688), PL-1 (EF199771) and PL-2 (EF199772) from Korea. The multiple sequence alignments were prepared with ClustalW and phylogenies were reconstructed with MEGA 4.1 software [17] using the Neighbor-joining tree inference method, with 1,000 bootstrap replicates to assign confidence levels to branches.
Chicken and turkey experiments
Experiment 1. Pathogenicity in immunosuppressed chickens
The pathogenic response of chemically immunosuppressed chickens to aMPV was studied. Thirty-nine 3 week-old specific pathogen free (SPF) chickens were divided into three groups of 13. Each group was housed in separated positive pressure isolators. Chickens in group 1 were inoculated intramuscularly with 100 mg/kg of CsA (Cypol; Chongkundang, Korea) per chicken, every 3 days from 3 weeks of age until the end of the experiment as previously described [18], and chickens in group 2 were not treated with CsA. At 31 days of age, both groups were inoculated oculonasally with 104 TCID50/0.1 mL of aMPV Ck/A/Kr/1336/06. Chickens in group 3 were kept as the untreated and unchallenged control. Nasal discharge and oropharyngeal swabs were collected from each group at 2, 3, 4, 5, 6 and 7 days post challenge (dpc). The virus titer of each specimen was determined using quantitative RRT-PCR. Standard curves were generated with control viral RNAs and the cycle threshold values of samples were converted into TCID50/mL by interpolation as previously described [19,28]. Three chickens from each group were sacrificed at 5 dpc and the turbinate, trachea and lungs collected for histopathological studies. Ten chickens of each group were monitored daily for 14 days for any clinical signs. Blood was collected from all remaining chickens and serum was tested for the presence of aMPV antibodies by enzyme-linked immunosorbant assay (ELISA). All experiments were carried out according to protocols approved by the Institutional Animal Care and Use Committee of Konkuk University.
Experiment 2. Pathogenicity in turkeys
Twenty-six 2 week-old turkeys were divided into two groups of 13. Each group was housed in separated positive pressure isolators. Turkeys in group 1 were inoculated oculonasally with 104 TCID50/0.1 mL of aMPV Ck/A/Kr/1336/06, while group 2 was the unchallenged control. Nasal discharge and oropharyngeal swabs were collected at 1, 3, 5, 7 and 9 dpc and tested for the shedding titer of aMPV by multiplex RRT-PCR. Three turkeys from each group were sacrificed at 5 dpc and the turbinate, trachea and lungs collected for histopathological studies. Ten turkeys of each group were monitored daily for 14 days for any clinical signs and blood collected to test for aMPV antibodies.
Histopathological examination
Nasal turbinate, trachea and lungs were collected from 3 chickens and turkeys at 5 dpc, formalin-fixed and processed. Paraffin sections were stained with hematoxylin and eosin using standard procedures for microscopic examination.
Serology
Sera were tested for the presence of aMPV antibodies using a commercial aMPV ELISA kit, avian rhinotracheitis antibody test kit (BioCheck, The Netherlands) according to the manufacturer's instructions. Based on the optical densities of the samples (405 nm), sample to positive (S/P) ratios were calculated and used to express the mean S/P ratio per group. Samples with an S/P of 0.5 or greater were considered to contain anti-aMPV antibodies, and were judged positive based on the manufacturer's recommendations. S/P of 0.35~0.499 was considered suspect for aMPV antibodies and a ratio of 0.349 or less was considered to be negative.
Results
Screening of the flocks by multiplex RRT-PCR
As shown in Table 2, four of the 156 samples from broiler chickens (2.6%), twenty of the 522 samples from broiler breeders (3.8%), and one of 102 samples from layers (1.0%) were detected by multiplex RRT-PCR. One subtype A and three subtype B aMPVs were detected from broiler flocks and 10 subtype A and 10 subtype B aMPVs were detected in broiler breeder flocks. One A subtype aMPV was detected in layer flocks.
Sequencing and phylogenetic analysis
Either 268 bp (subtype A) or 361 bp (subtype B) fragments of the G gene determined from all aMPV positive samples by multiplex RRT-PCR were amplified by RT-PCR for sequence analysis. A yield of seven partial sequences of the G gene was the result. The nucleotide sequences of the G gene of aMPV subtypes A and B received the following Genbank accession numbers: Ck/A/Kr/567/04 (FJ796704), Ck/A/Kr/166/04 (FJ796707), Ck/A/Kr/167/04 (FJ796706), Ck/Kr/A/1336/06 (FJ796703), Ck/A/Kr/301/08 (FJ796705), Ck/B/Kr/168/04 (FJ796702) and Ck/B/Kr/746/04 (FJ796701). Five genes clustered with subtype A reference strains and two genes clustered with subtype B reference strains (Fig. 1).
Virus isolation in Vero cells
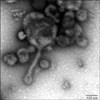
To isolate aMPV from field samples, IBV, NDV, ILTV and Mycoplasma gallisepticum were also screened. Samples that were positive for any of these other pathogens were not used for aMPV isolation, and the samples that were RRT-PCR positive for aMPV were candidates for virus isolation. The infected cell layer of each passage was trypsinized and reseeded into culture flasks. CPE was characterized by many rounded and floating cells, and appeared in Vero cells at 3~4 passages. Unfortunately, 23 aMPV positive samples by multiplex RRT-PCR disappeared upon passaging, and just two subtype A aMPVs (Ck/Kr/A/1336/06 and Ck/A/Kr/301/08) were isolated in Vero cells (Table 2). Ck/Kr/A/1336/06 was isolated from a broiler breeder and Ck/A/Kr/301/08 was isolated from a layer. When examined by negative stain electron microscopy, Ck/A/Kr/1336/06 appeared as long filamentous forms (Fig. 2). In conjunction with electron microscopy, the isolate was confirmed by immunofluorescence assay using TRT12 monoclonal antibody (data not shown). CK/Kr/A/1336/06 isolated by passaging 4 times was directly used for pathogenicity testing in chickens and turkeys.
Experimental infection in SPF chickens and turkeys
Experiment 1
Because SHS cannot be induced by aMPV alone in chickens [10], SPF chickens were immunosupressed by CsA followed by inoculation with Korean aMPV to reproduce the disease. Both CsA-treated and non-treated chickens showed only nasal discharge at 3 dpc and did not show any histopathological lesions. None of the non-challenged chickens showed clinical signs or histological lesions. Viral replication was detected in the nasal discharge of the infected birds by RRT-PCR. As shown in Fig. 3, there were no differences in viral load between CsA-treated and non-treated chickens at 2, 3 and 4 dpc. However, at 5 (p < 0.05), 6 (p < 0.05) and 7 (p < 0.005) dpc, CsA-treated chickens excreted viruses at a higher titer than non-treated chickens. Although virus shedding from nasal discharge had ceased by 7 days post infection in non-treated chickens, CsA-treated chickens continuously shed the virus until 7 days post infection.
Experiment 2
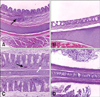
To determine the pathogenicity of Korean aMPV in commercial turkeys, 10 two-week-old turkeys were inoculated with Korean aMPV. Four of the 10 turkeys began to show nasal discharge at 3 dpc, and seven of the 10 turkeys showed symptoms such as nasal discharge, sinus swelling and watery eyes at 5 dpc (data not shown). None of the non-challenged turkeys showed clinical signs or histological lesions. The presence of clinical signs in aMPV infected turkeys was associated with histological lesions characterized by mild to severe inflammation in the turbinate and trachea (Fig. 4). As shown in Fig. 3, significantly higher virus titer was shed from nasal discharge than oropharyngeal swabs at 3 (p < 0.05), 5 (p < 0.05), and 7 (p < 0.005) dpc, and aMPV infected turkeys shed the virus from both oropharyngeal swab and nasal discharge until 9 dpc.
Serology
On the day of aMPV challenge, all sera collected from birds confirmed that chickens and turkeys were negative for aMPV (Table 3). In experiment 1, at 14 dpc, three of 10 sera of CsA-treated SPF chickens were positive for aMPV antibodies (mean S/P ratio = 0.58 ± 0.55). In CsA non- treated chickens, two of 10 sera were suspect for aMPV antibodies. No antibodies against aMPV were detected in serum samples of CsA non-treated and non-challenged chickens (0.10 ± 0.06). In experiment 2, seven of 10 sera of turkeys were positive, showing high antibody levels (mean S/P ratio = 1.03 ± 0.85) and no antibodies against aMPV were detected in serum samples of non-challenged turkeys (0.10 ± 0.01) at 14 dpc.
Discussion
Although SHS has been reported in Korea since 1992 [16], this is the first study isolating and characterizing aMPV subtype A from chickens in Korea. In this study, we isolated two subtype A aMPV strains from chickens. One was isolated from a broiler breeder flock showing facial swelling especially around the eye, nasal discharge, egg production losses and discolored eggs. The other subtype was isolated from a layer flock in which egg production losses were evident (data not shown). According to necropsy results of broiler breeders, severe nasal discharge induced blockage of the larynx of the chickens, delaying the feeding time, resulting in a fatty liver and, finally, lower productivity. In most reports of aMPV infection in chickens, the focus is on SHS associated with aMPV. In this study, however, we assumed that severe nasal discharge induced by aMPV caused delayed feeding time and a lower productivity, as well as SHS linked with aMPV.
Due to the poor replication of aMPV in chickens [8,12], isolation of aMPV from chickens is limited. Furthermore, because clinical signs induced by aMPV alone are not apparent in chickens, virus isolation from chickens is more difficult than from turkeys [12]. We determined that three factors are important in the isolation of aMPV from chickens. The first one is sampling at the early stage of the aMPV infection. Sampling in the early stage of the infection is crucial, as the virus may be present only in the nasal sinuses and turbinates for a short period when the chicken showing the very first symptoms of the disease [7]. The second factor concerns the type of sample. We attempted to isolate the aMPV from the nasal turbinate of chickens that did not show clinical signs in broiler breeder flock showing SHS, and ascertained that the detection rate of aMPV was higher in nasal turbinates than in oropharyngeal swabs (data not shown). The third important factor concerns the isolation method; reseeding of the aMPV by passage. Lu et al. [22] isolated the aMPV strain LU using tracheal organ culture and adapted aMPV to Vero cells passaged by trypsinizing and reseeding into culture flasks. In a slight modification to isolate aMPV from field samples, we directly inoculated these samples into Vero cells and passaged by trypsinizing and reseeding into culture flasks. Because of the poor replication of aMPV in the host [8,12], aMPV titer may be very low in the early passages of Vero cells. Therefore, it appears to be very critical that host cells be passaged by trypsinizing and reseeding into culture flasks.
As reported in Japan and Taiwan [22,23], and due to our geographic location, we expected to diagnose aMPV subtypes A and/or B at a high rate. Indeed, the survey results of aMPV from chicken flocks showed aMPV infection with co-circulation of subtypes A and B. Sequence analysis of parts of the attachment protein G gene showed a relative homogeneity within the Korean aMPV genes and confirmed that the aMPV detected samples belonged to subtypes A and B.
In animal experiments, although we induced immunosuppression in chickens to reproduce SHS, CsA treatment of chickens could not reproduce SHS. A previous study also unsuccessfully attempted to induce re-excretion of aMPV in poults and chicks using CsA immunosuppression [15]. The failure might reflect a situation where reproduction of SHS is not under T-cell control. However, considering the antibody responses in CsA-treated chickens in this study, it seems that T-cell suppression rendered the chickens more susceptible to infection by a aMPV isolate than non- CsA treated chickens. Although aMPV shedding titer from the nasal discharge of chickens and turkeys was similar, turkeys only showed clinical signs and histopathological lesions. These data demonstrated that the Korean aMPV isolate also causes severe respiratory infection in turkeys than chickens, as previously reported [6]. In contrast to the turkeys, aMPV infected chickens showed only transparent nasal discharge in spite of CsA treatment. Rhinovirus causes common cold accompanied with nasal discharge without the damage of nasal epithelium in humans. And nasal obstruction can interfere with sleep and feeding [30]. Despite the presence of mild clinical signs and virus shedding, histological lesions were not observed in affected chickens. Although further studies are essential, the pathogenesis of aMPV in chickens may be similar to rhinovirus infection in humans. As rhinovirus infected humans have disruptions in feeding, we could also check the delayed feeding time in aMPV infected broiler breeders caused by obstruction of the larynx. Consistent with a previous report [25], aMPV infected chickens also showed very low serum antibody levels. Because the aMPV replicate in upper respiratory tissues for only a short time [9], the low multiplication levels of aMPV in target tissues of chickens results in low antibody responses. These results could explain why the re-infection of aMPV occurs frequently in chicken flocks.
In summary, we confirmed aMPV infection in chickens and isolated aMPV from chickens in Korea. aMPV isolation from chickens showing severe nasal discharge suggests that it induces lower productivity due to upper respiratory passage blockage in chicken flocks. These results suggest aMPV infection in chicken flocks gives rise to appreciable economic losses in the poultry industry. Therefore, continuing epidemiological surveys are necessary for the prevention of aMPV infections in chicken flocks in Korea.





 XML Download
XML Download