

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Ulceration in the non-glandular part of the stomach of pigs has been reported in several countries [12,30,31]. It is a disease of complex etiology in which multiple factors are involved including dietary and stress conditions [19] such as small particle sizes of feed, interruption of feed intake and the presence of highly fermentable carbohydrates [1]. Its etiology also seems to be associated with various infectious diseases such as porcine reproductive respiratory syndrome (PRRS), post-weaning multisystemic wasting syndrome (PMWS) and porcine dermatitis nephrophathy syndrome (PDNS) [14,32]. Ulcers of the non-glandular part of the stomach have been associated with Helicobacter (H.) suis [4,5,28,31], but the exact role of H. suis in porcine gastric pathology is a subject of considerable debate. Indeed, several authors [11,23,26,33] did not find this association while it has been recently reported that gastric ulcers of the non-glandular part of the stomach were induced in pigs experimentally infected with H. suis [13]. In any case, it was demonstrated that H. suis causes gastritis in experimentally and naturally infected pigs [3,11,18,24,26,28]. H. suis is the main Helicobacter species colonizing the stomachs of pigs and is the most prevalent gastric non-H. pylori Helicobacter species in humans [35]. There are strong indications that pigs may be a source of H. suis infection in humans, while animals may be occasionally infected with H. pylori [7,9,34]; Krakowka et al. [20] demonstrated the occurrence of gastric ulcers in gnotobiotic piglets experimentally infected with H. pylori. Our study was conducted to evaluate the presence of Helicobacter spp., H. suis and H. pylori by PCR in swine affected by gastric ulcers. Moreover, the presence of Helicobacter spp. was investigated in feces and saliva samples in order to assess the feasibility of an in vivo detection method.
Materials and Methods
Samples
A total of 400 stomachs collected at slaughter from 10-month-old pigs (50% males and 50% females) coming from two intensive closed-cycle-based breeding farms in the centre of Italy, were examined to evaluate the presence of gastric ulcers. For molecular studies, feces and saliva swabs were also collected from 25 subjects belonging to the same groups and stored at -80℃ until use. Each stomach was individually examined using disposable blades and gloves to avoid cross contamination. The stomachs were opened longitudinally along the greater curvature. The gastric content was gently removed and the mucosa was examined to evaluate the presence of pathological changes. Sixty-five samples from the pars esophagea were taken from pigs with ulcers and 15 samples were collected from pyloric portions without macroscopic signs of ulcers. All these samples were cut and divided into two parts and then stored in an ice box (4℃) for transport to the laboratory for histopathology and molecular analyses.
Histopathology
The samples were fixed in 10% phosphate-buffered formalin, dehydrated, embedded in paraffin, sectioned at 4 µm, and stained with hematoxylin and eosin for light microscopic examination.
DNA extraction and polymerase chain reaction (PCR)
All the samples were stored at -80℃ until use. The DNA was extracted from all the samples using the Charge Switch gDNA mini tissue kit (Invitrogen, UK) in accordance with the manufacturer's protocol. For PCR analysis, specific primers (Invitrogen, UK) and PCR protocols were used (Table 1) and the genomic DNA of H. pylori (ATCC 43504) was employed as a positive control for Helicobacter spp. and H. pylori PCRs. Reactions were performed in a 50-µL volume consisting of 1.25 µg of extracted DNA, 2 U of Taq DNA polymerase, 0.5 µM of each primer, 1.5 mM (2.5 mM for H. suis PCR) of MgCl2, 200 µM of dNTPs mix and ×1 PCR buffer (Invitrogen, UK) in a Gene Amp PCR System 2400 thermal cycler (Applied Biosystems, USA), according to the specific PCRs conditions (Table 1). PCR products were resolved by electrophoresis in 1~1.5% agarose gel containing 4 µL of GelRed (nucleic acid gel stain, ×10,000 in water) per 100 mL (Biotium, USA) and examined by transillumination (Euroclone, Italy).
Restriction enzyme digestion
To support H. suis PCR analysis, 10-µL samples, of Helicobacter spp. PCR product were digested with 10 U of MboI (Gibco, UK) for 4 h at 37℃ [27]. The digested samples were analyzed by electrophoresis in 1.5% agarose gel and examined by transillumination.
DNA sequencing
Eight H. suis PCR products from the gastric samples were sequenced using V832f primer, specific to the H. suis 16S rRNA gene, and 8 Helicobacter spp. PCR products from feces and saliva samples were sequenced using H276f primer, specific to the 16S rRNA gene of Helicobacter genus. All samples for nucleotide sequencing were obtained by PCR product purification using the Jet Quick gel extraction spin kit (Genomed GmbH, Germany) in accordance with the manufacturer's protocol. PCR fragments and primers were premixed in the same tube, and sent to the company (Primm Srl, Italy), for sequencing. The DNA sequences obtained were aligned and compared with the BLAST database (NCBI, USA).
Results
Gross lesions and histopathology
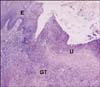
Three hundred and seventy-three of the 400 stomachs (93%) examined at the slaughterhouse showed ulcers across the small curvature of the pars esophagea, often associated with extensive hyperemic areas. At times, the edges of the ulcers appeared thickened and firm. Histological examinations frequently revealed lymphocytic and plasmacytic cellular infiltrates in the glandular part in acute evolution and lymphoid follicle formations in the gastric lamina propria; instead lymphofollicular gastritis was predominant in chronic ulcerous processes (Fig. 1). Proliferation of granulation tissue and hyperplasia of the epithelium were also visible near ulcers in sub acute and chronic evolution.
PCR analysis
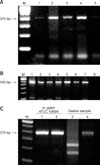
PCR analysis of the gastric samples revealed the presence of Helicobacter spp. in 30/65 (49%) of the esophago-gastric ulcer samples and 8/15 (53%) of the samples from pyloric mucosa without lesions. Moreover, all Helicobacter spp. positive samples were also positive for H. suis PCR (Figs. 2A and B). H. pylori was not detected in any samples. PCR analysis of feces and saliva samples showed that 15/25 (60%) and 16/25 (64%) were positive for Helicobacter spp., respectively. None of the samples was positive for H. suis or H. pylori.
Restriction enzyme analysis
As expected, MboI cleaved the 376 bp Helicobacter spp. PCR products, amplified from the gastric samples, into two fragments of 147 and 229 bp, with the exception of those from H. pylori (ATCC 43504) used as negative controls (Fig. 2C).
DNA sequencing
Sequence analysis from 8 gastric samples positive for H. suis PCR showed 99% homology with the 16S rRNA gene of H. suis [2]. Sequence analysis from 8 feces samples and from 8 saliva samples positive for Helicobacter spp. showed 99% homology with the 16S rRNA gene of Helicobacter genus.
Discussion
Ulceration of the non-glandular esophageal portion of the stomach of feeder pigs, known as gastroesophageal ulceration, is a common and serious problem in swine production [10]. Its etiology is multifactorial, including genetic, nutritional, environmental and infectious causes; indeed, animals affected by PRRS, PMWS and PDNS usually show gastric lesions at necropsy, but the relationship seems to be circumstantial [14,19]. Our results showed that ulcers of the non-glandular esophageal portion of the stomach were present in a high percentage of the slaughtered pigs (93%), as confirmed by histological examination. Ulcers were not macroscopically observed in the glandular part (pyloric portion), while histological examination frequently showed lymphocytic and plasmacytic cellular infiltrates, as reported elsewhere [17]. PCR analysis of gastric samples from the pars esophagea (with ulcers) and from the pyloric portion (without ulcers) revealed the presence of Helicobacter spp. in 49% and 53% specimens respectively. Moyaert et al. [25] compared different Helicobacter spp. PCR primers showing that the ones described by Riley et al. [29] and used in this study are highly capable of correctly identifying Helicobacter strains and have a moderate ability to exclude non target species, giving amplicons of the correct size. To determine if the microorganisms present in the gastric samples were H. suis or H. pylori, the Helicobacter spp. positive samples were submitted to H. suis and H. pylori specific PCR. All the samples were positive for H. suis and negative for H. pylori. The results for H. suis were confirmed by restriction enzyme analysis and DNA sequencing. Our results may indicate that H. suis colonizes both the pars esophagea [31] and the pyloric portion with ulcer occurrence only in the pars esophagea. The presence of ulcers in the pars esophagea suggests a different susceptibility of the two anatomic regions to the same pathogen noxa. This may be due to their peculiar morphology; the stratified squamous epithelium of the pars esophagea is devoid of mucous-producing glands and lacks the sodium bicarbonate buffering system present in the gastric glandular mucosa, thus favouring the occurrence of damage caused by gastric acid content [2,7,13]. Indeed, it has been reported that H. suis mainly colonizes the antrum and the fundic gland zone and, to a lesser extent, the cardiac gland zone [4,13,17]. In the fundic gland region of pigs experimentally or naturally infected with H. suis, these microorganisms were found in close contact with parietal cells, which indicates that the bacterium may have an impact on hydrochloric acid-producing cells [13,17]. The secretion of excessive amounts of gastric acid may lead to increased contact of the non-glandular part of the stomach with hydrochloric acid. Consequently, this chronic insult of the non-glandular part may result in the development of ulcers. The porcine stomach physiologically maintains the proximal and the distal compartments with distinct pH and enzymatic conditions [13].
Our study did not detect H. pylori in the samples analyzed by PCR. Although Krakowka et al. [20] reported that H. pylori is able to colonize the gastric mucosa of experimentally infected pigs, there are no indications that swine are a natural reservoir of this pathogen [8,13]. To assess an in vivo detection method for H. suis, PCR analyses with specific primers were carried out on saliva and feces samples. In our study 60% and 64% of feces and saliva samples were positive in the Helicobacter spp.. PCR respectively, while they were negative for H. suis and H. pylori DNA. Sequence analysis from 8 positive samples of feces and from 8 positive samples of saliva showed 99% homology with Helicobacter spp.. Our data are in accordance with the results of other authors. Indeed, Hänninen et al. [15,16] isolated H. bilis and H. trogontum from feces samples while H. suis has never been detected.
In conclusion our results show that although PCR is effective for the detection of H. suis in gastric samples, it might be less sensitive in saliva and feces samples. There are indications that Helicobacter spp. different from H. suis, may be present in feces and saliva.

 XML Download
XML Download