

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Tick-borne encephalitis virus (TBEV) is a member of flaviviridae, genus flavivirus, that causes inflammatory infections of the central nervous system presenting as meningitis, meningoencephalitis, or meningoencephalomyelitis in European countries, northern Russia, China, and Japan [2,10,14]. TBEV is subdivided into three subtypes; Far Eastern [Russian spring-summer encephalitis (RSSE)], Western [Central European encephalitis (CEE)], and Siberian that demonstrate various severities of disease [7]. RSSE infections frequently result in very severe encephalitis with mortality rates ranging from 20~60%, while CEE infections caused by the Western subtype induce milder symptoms with mortality rates of 1~5% [7]. Ixodes (I.) ricinus is the primary vector of the Western subtype, while I. persulcatus is the primary vector of the Far Eastern and Siberian subtypes [7,15].
In Korea, Haemaphysalis spp. are the predominant ticks collected in tick drag surveys, while I. nipponensis is the primary species collected from small mammals [20]. The protein E gene of the Western subtype of TBEV was detected by RT-nested PCR and TBEV was isolated from lung and spleen tissues of Apodemus agrarius Thomas, the striped field mouse, captured in northern Gangwon-do and Gyeonggi-do provinces, Korea, and from Haemaphysalis (H.) longicornis and I. nipponensis ticks, which previously had not been reported as TBEV vectors [5,11]. Only the Far Eastern subtype was detected in countries neighboring Korea, including Japan, China, and northeastern Russia [5,14]. The purpose of our investigation was to evaluate the potential for TBEV occurrence in Jeju Special Self-Govering Province (Jeju Island) and southern Korea.
Materials and Methods
Tick collections
Ticks surveys were conducted using tick drags (1 × 1 m white cotton flannel cloth) through various vegetative habitats, including deciduous, pine, and mixed forest leaf-litter, grasses, and herbaceous vegetation, in 2007 at Jeju Island, Jeollanam-do, Gyeongsangbuk-do, and Gyeongsangnam-do provinces. Ticks were placed individually into 1.5 µL cryovials, labeled with a collection site number, transported to the laboratory, and identified to species and development stages under a dissecting microscope [30]. Identified ticks (total 6,788 larvae, nymphs, and adults) were returned to the original vial with a unique identification number and stored at -70℃ until assayed.
RT-nested PCR
A total of 4,077 (60.1%) of 6,788 ticks collected were pooled according to species, developmental stages, and collection sites into 649 pools (larvae, 1~14; nymphs, 1~20; adults, 1~5 ticks/pool), and homogenized using beadbeater, Tissuelyser II (Qiagen, Germany), 30 cycles/sec with a tissue lysis buffer and 5 mm stainless steel beads, and then centrifuged at 12,000 g for 15 min at room temperature (Table 1). After centrifugation, the supernatant was used for RNA extraction.
Total RNA was extracted from the tick suspension using RNeasy mini kit (Qiagen, Germany) according to the manufacturer's instructions. The RNA was eluted with 40 µL of distilled water and stored at -80℃ until used. The cDNA synthesis was performed using Sensiscript RT kit (Qiagen, Germany) according to the manufacturer's instructions.
For detection of the TBEV envelope glycoprotein gene, primary reactions were carried out with i-Star Master mix PCR kit (Intron, Korea) and previously designed primers, TBE913F (5'-TGCACACAYYTGGAAAACAGGGA-3') and TBE1738R (5'-TGGCCACTTTTCAGGTGGTACTTGGTTCC-3') [27]. RT-nested PCR was conducted in a PCR thermal cycler (PTC-200, MJ Research, USA) and the primary reaction was performed with an initial denaturation of 5 min at 94℃, followed by 30 cycles of 30 sec at 94℃, 1 min at 53℃, and 1 min at 72℃, with a final extension step of 5 min at 72℃. Two µL of synthesized cDNA and 18 µL of i-Star Master mix solution (Intron, Korea) was used in a total volume of 20 µL. Nested PCR was carried out with an i-Star Master mix PCR kit (Intron, Korea) of 30 cycles for 20 sec at 94℃, 10 sec at 65℃, 20 sec at 68℃, with a final extension step of 5 min at 72℃. The nested reaction was conducted using 1 µL of the primary PCR product as a template and previously described primers [TBE1192F (5'-CAGAGTGATCGAGGCTGGGGYAA-3') and TBE1669R (5'-AACACTCCAGTCTGGTCTCCRAGGTTGTA-3')] [29]. Samples that were found to be positive by previous screening were selected for a second PCR assay with the following NS5-gene specific primers: outer forward primer, 5'-GAGGCTGAACAACTGCACG-3'; outer reverse primer, 5'-GAACACGTCCATTCCTGATC T-3'; internal forward primer, 5'-ACGGAACGTGACAAGGCTAG-3'; and internal reverse primer, 5'-GCTTGTTACCATCTTTGGAG-3' [17]. Thirty-five amplification cycles were performed, with each cycle consisting of 30 sec at 96℃, 30 sec at 40℃, and 30 sec at 68℃, with 1.0 µL of this amplification mixture subjected to 30 cycles as above of the nested PCR with the internal primers. The PCR products were analyzed by agarose gel electrophoresis, with a 100 bp ladder molecular weight DNA size marker (Intron, Korea) in 1.5% agarose gel, stained with ethidium bromide.
Cloning, sequencing, and phylogenetic analysis
To confirm positive TBEV PCR products, all purified positive amplicons were ligated and cloned in the pGEM-T Easy Vector system (Promega, USA). The vectors were transformed in thermo-competent Escherichia coli DH5α cells. The plasmid DNAs were amplified and purified with DNA-spin plasmid DNA purification kit (Intron, Korea) and verified by sequence analysis. The cloned DNA was sequenced by a cycle sequencer using ABI Prism 3700 DNA analyzer (Applied Biosystems, USA). Sequence homology searches were made at the National Center for Biotechnology Information (NCBI, USA) BLAST network service. Comparative analysis of the amino acid sequences (protein E gene) and nucleotide sequences (NS5 gene) was completed using TBEV sequences in the GenBank database. Phylogenetic analysis was completed using the ClustalX 1.60 program [28] and a phylogenetic tree deduced by neighbor-joining method PAUP 4.0b software for Macintosh (Sinauer Associates, USA). Numbers on the branches represent bootstrap support (more than 50%) generated from 400 replications.
Sequences were obtained from GenBank database. Accession numbers for TBEV protein E gene partial amino acid sequences are as follows: AB062063 (TBEV strain Oshima); AY182009 (TBEV strain Senzhang); AF091019 (TBEV strain T-Blood); AB062064 (TBEV strain Sofjin); AY217093 (TBEV strain MDJ-01); AF091008 (TBEV strain Crimea); AF091013 (TBEV strain N132); AFO91016 (TBEV strain RK1424); L40361 (TBEV strain Vasilchenko); AF091006 (TBEV strain Aina); AF527415 (TBEV strain Zausaev); EU276110 (TBEV strain Krm213); EU276113 (TBEV strain Krm219-1); EU276111 (TBEV strain Krm215); DQ988684 (TBEV strain Krm219); EU276109 (TBEV strain Krm93); AF091010 (TBEV strain K23); U39292 (TBEV strain Hypr); DQ992536 (TBEV strain Krm216); AF091007 (TBEV strain Als.I) NC005062 (Omsk hemorrhagic fever virus strain Bogoluvovska); NC003687 (Powassan virus strain LB).
Accession numbers for NS5 gene sequenes are as follows: U39292 (TBEV strain Hypr); FJ572210 (TBEV strain Salem); AM600965 (TBEV strain K23); FJ159003 (TBEV strain FVG-ML); AB062063 (TBEV strain Oshima); AB062064 (TBEV strain Sofjin); FJ906622 (TBEV strain Primirye-89); AY182009 (TBEV strain Senzhang); AY217093 (TBEV strain MDJ-01); DQ989336 (TBEV strain 205); L40361 (TBEV strain Vasilchenko); DQ451301 (TBEV strain Kokkola-26); NC005062 (Omsk hemorrhagic fever virus strain Bogoluvovska ); NC003687 (Powassan virus strain LB).
Results
Tick identifications
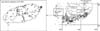
A total of 6,788 ticks (78 larvae, 6,444 nymphs, and 266 adults) were collected from 113 collection sites of various types of habitats on Jeju Island, and the southern tip of the Korean Peninsula and associated islands, were identified to species (Fig. 1). In total, 4,077 ticks (39 larvae, 3,805 nymphs, and 233 adults) were pooled and assayed for TBEV. H. longicornis was the most frequently collected (73.4%; n = 4,984 [2,917 tested]), followed by H. flava Neumann (22.4%; n = 1,523 [929 tested]), H. phasiana Saito, Hoogstraal, and Wassef (3.2%; n = 216 [167 tested]), Amblyomma testudinarium C. L. Koch (0.4%; n = 25 [25 tested]), I. nipponensis (0.4%; n = 24 [24 tested]), and I. turdus Nakatsuji (0.2%; n = 16 [16 tested]) (Table 1, Fig. 1).
Detection of TBEV from ticks
TBEV was detected in two of the six collected species (H. longicornis and H. flava) (Table 1). The minimum field detection rates (assumes a minimum of one infected tick/pool) of TBEV were 0.17% for H. longicornis and 0.14% for H. flava. Using PCR primer sets targeting the protein E gene of TBEV, 6/649 pooled tick samples of 4,077 ticks were PCR positive, which produced a 477 bp amplicon. All six of the positive PCR pools were collected from Jeju Island, while no positive amplicons were detected from other areas surveyed (Table 2, Fig. 1).
Sequence analysis and phylogenetic tree
The amplified 477 bp region of the protein E gene and 252 bp region of the NS5 gene of six positive samples at five collection sites were sequenced and compared with other TBEV sequences of northeast Asia available in the GenBank database. The NS5 gene sequences are presented in Fig. 2. Acquired NS5 gene sequences showed a 98% identity to the Western subtype K293 (Czech) strain, 81.7% identity to the Siberian subtype strain Vasilchenko (Russia) and 82.5% identity to the Far Eastern subtype strain Oshima (Japan). The phylogenetic trees of the protein E genes with support positions are shown in Fig. 3. Positive amplicons of the protein E gene (GenBank accession number FJ972625) from Jeju Island belong to the Western subtype (Fig. 3). Strains Als.I (AF091007), Aina (AF091006), and Oshima (AB001026), are representative strains for Western, Siberian, and Fareastern subtypes, isolated from France, Russia, and Japan, respectively [7,24]. The protein E gene nucleotide sequences of positive amplicons demonstrated a closer homology with the Western subtype (99.4%) than with the Siberian (97.6%) and Far Eastern (97.6%) subtypes.
Discussion
Since 2000, noticeable increases have been seen in the number of tick-borne encephalitis (TBE) cases in Europe and in the number of isolations of TBEV from wild rodents and ticks in endemic countries, e.g., Czech Republic, Germany, Slovenia, Sweden, Switzerland, and Korea [12,22,23]. Neighboring countries, including Japan, China, and Russia have reported a high incidence of TBEV infections, with Russia having the highest number of reported Siberian and Far Eastern TBEV cases [4,13,23]. Only the Far Eastern TBEV subtype has been reported from China and Japan [7,14]. While northern Japan is highly endemic for TBEV with numerous reports of transmission to sentinel animals and isolations from ticks, few human cases have been reported. In China [14,25,26], the first human TBE case was detected in 1943 and the Far Eastern TBEV subtype is mostly reported from the northeastern forests and mountains of Changbai, Jinlin province, Daxing'an, Inner Mongolia province, and Xiaoxing'an, Hei Longjiang province, which extend into North Korea. I. persulcatus is the primary vector of the Far Eastern TBEV subtype of Japan and China [1,14]. Outbreaks of human cases of TBEV and its geographical distribution of the Western and Far Eastern subtypes largely follow that of their primary tick vector distributions, I. ricinus and I. persulcatus, respectively [16].
While it would seem reasonable that the Far Eastern TBEV subtype would be present in Korea, only the Western subtype has thus far been isolated from A. agrarius [12], while the Western subtype TBEV protein E gene was identified from H. longicornis, H. japonica, H. flava, and I. nipponensis [11]. Based on these findings, we presume that there is a high probability of the coexistence of the Western and Far Eastern TBEV subtypes in Korea. Only limited surveys of TBEV have been conducted in Korea, primarily in Gyeonggi-do and Gangwon-do provinces, and more comprehensive surveys where there are relatively high populations of I. persulcatus may result in the identification of coexisting TBEV subtypes.
Globally, ten species of ticks (I. ricinus, I. persulcatus, I. hexagonus, I. arboricola, I. ovatus, H. punctata, H. concinna, H. inermis, Dermacentor marginatus, and D. reticulatus) are reported to be vectors of TBEV [8,20,21]. Currently, five species incriminated as TBEV vectors (H. longicornis, H. japonica, H. flava, I. persulcatus, and I. nipponensis) have been reported in Korea [11,12]. Of the six species assayed for TBEV in our study, only H. longicornis (five pools) and H. flava (one pool), the most frequently collected ticks in grass and forested habitats, respectively, were positive by PCR for TBEV. Although TBEV infected ticks have been reported on mainland Korea [11,12], in our study only H. longicornis and H. flava collected from Jeju Island were positive by PCR for TBEV. The distribution and population densities of tick vectors are considered important elements in the distribution and prevalence of each of the TBEV subtypes [11]. Furthermore, range expansion of TBEV vectors due to world climate and ecological changes are regarded as key factors of the recent expansion of TBEV [1,3,18,19]. In Korea, as a result of a tree planting policy established in the 1960s, previously relatively barren hills and mountains are now covered with dense young deciduous and conifer forests. This has dramatically altered the ecology of these areas, creating the potential for increases and variation of animal populations and their associated ectoparasites, which are dependent upon forested areas for refuge. Additionally, migratory birds, considered as primary hosts for many tick species, are responsible for transporting TBEV and their vectors [6,9]. Recently, TBEV was identified from ticks infesting migratory birds in Sweden, suggesting that birds may be a competent reservoir by serving as a host for all stages of potential tick vectors that infest wild birds [29].
Large populations of migratory birds visit Jeju Island annually, possibly transporting ticks or infecting indigenous ticks that feed on potentially infected migratory birds during their stay. Based on Chinese data, both the Far Eastern and Western subtypes are likely to coexist in North Korea [14]. More comprehensive surveys throughout Korea are required to determine the presence, geographical distributions, and prevalence of TBEV subtypes in Korea.
Furthermore, for other Asian countries where undiagnosed encephalitis outbreaks occur and I. persulcatus and I. ricinus are not primary tick species, investigations to determine the source of the infections are needed. Our results add to the growing evidence that other species of ticks, including H. longicornis and H. flava, serve as potential vectors of the TBEV Western subtype.




 XML Download
XML Download