

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
The influenza virus is a member of the orthomyxoviridae family of RNA viruses, and has three genera - A, B and C. Each genus of the virus is further subdivided by serotype based upon the surface viral proteins hemagglutinin (HA) and neuraminidase (NA). Avian influenza viruses of the Influenza A group of viruses only infect birds. Currently, 16 HA and 9 NA virus subtypes have been described in the avian species, and avian influenza viruses with various subtype combinations have been isolated [1,2].
Diagnosing infection with avian influenza virus (AIV) can be accomplished in two ways: either through direct detection of the virus itself, or through detection of antibodies specific for the virus. Direct detection is accomplished by isolation of the virion particles, by viral antigen detection through immunospecific assays, or by targeting viral-specific nucleic acids in the serum of suspicious animals. The hemagglutinating inhibition (HI), agar gel precipitation (AGP), and virus neutralization tests are common serologic tests for AIV employed in the diagnostic laboratory [4,7,11,12].
As the AIVs become widely distributed throughout the world, there is an increased need for AIV surveillance and tests for AIV vaccine efficacy. The HI test is inexpensive, and is therefore the most commonly used test to determine the presence of antibodies in the serum to an influenza virus [7,13]. However, the HI test requires several standardizations and is labor-intensive. Additionally, reference sera always need to be included when performing the test since minor variations in the epitopes of the viral subtypes can result in inaccurate results [12].
The enzyme-linked immunosorbent assay (ELISA) is another option for AIV surveillance and/or evaluation of vaccine efficacy. ELISA methods are subdivided based on their approach, and include both indirect and competitive methods. The main benefit of the competitive ELISAs (cELISAs) over indirect ELISAs in AIV surveillance is that indirect ELISAs require the use of species-specific enzyme-conjugated antibodies, while cELISAs do not. The use of cELISA \eases the testing of sera from various species. Several ELISA methods have been developed to detect AIV antibodies in animal sera, all of which target the common viral proteins of Influenza A viruses [6,8,14,17-19]. In addition, previous studies have suggested that cELISAs should be effective for large-scale surveillance of AIV in avian flocks or herds of other species [17].
In this study, we addressed whether a specific, commercially-available cELISA could be specific and sensitive enough for seroepidemiologic or vaccine efficacy studies. The seroreactivity of the cELISA were evaluated against sera from both avian and non-avian species.
Materials and Methods
Reference sera and viruses
The reference sera for AIV subtypes, Newcastle Disease virus (NDV), Mycoplasma (M.) gallisepticum, and Egg drop syndrome (EDS)-76 virus were provided by the OIE AIV Reference Laboratories that validated serum reactivity: Instituto Zooprofilattico Sperimentale (IZS) delle Venezie, Italy and the Federal Center for Animal Health (ARRIAH), Russia. Negative sera from specific pathogen free (SPF) chickens were used for the validation of this ELISA in this experiment (ND SPF, Korea).
The virulent AIV H9N2 strain was provided by the National Veterinary Research and Quarantine Service (Korea) for the seroconversion test after viral challenge. For the checking of seroconversion in seum after influenza virus vaccination via intramuscular route, H9N2 and H5N1 vaccine strains from the Research Unit of Green Cross Veterinary Products (GCVP), Korea and ARRIAH of Russia, respectively, were used.
Serologic tests
HI and AGP tests were performed according to OIE manuals. The overall protocol for the single radial hemolysis (SRH) test was performed according with guidelines of the Manual of Diagnostic Tests and Vaccines for Terrestrial Animals (Equine influenza, part 2-section 2.5-chapter 2.5.5). The commercially-available cELISA evaluated in this study was manufactured by Animal Genetics (Korea), and was performed according to the manufacturer's instructions. Briefly, the cELISA plate, pre-coated with the viral nucleoprotein (NP) antigen, was incubated with an equal mixture of sample and monoclonal (anti-NP) HRP-conjugated antibodies for 30 min at 37℃. Following this incubation, all unbound material was removed by aspiration and the wells were washed prior to addition of a substrate solution. The residual enzyme activity in each well was found to be inversely proportional to the concentration of anti-NP antibodies in the specimen. The results were expressed as the percent inhibition (PI) value according to the following formula:
PI value = [1-(OD sample/mean OD negative)] × 100
Based on the PI value and animal species, the samples were classified as positive if the PI value was above 50 (except for turkey, which was considered to be positive for PI values > 85), and negative if the PI value was below 49 (with the exception of turkey, which was negative for PI values < 84).
A second competitive ELISA (IZS ELISA) was used to compare the specificity and sensitivity of the AG cELISA with another cELISA method. The IZS ELISA was performed according to the manufacturer's instructions.
Indirect ELISA methods, including ProFLOCK Plus AIV Ab test kit (Synbiotics, USA), Avivac (ARRIAH, Russia), and FlockChek Avian Influenza Antibody Test Kit (IDEXX, USA), were performed to compare indirect and competitive ELISAs. The indirect ELISA methods were performed following the manufacturers' instructions.
Seroreactivity test with reference sera
Seroreactivity tests were performed by two OIE AIV reference laboratories, IZS and ARRIAH. H1 to H15 AIV subtypes, NDV, EDS-76 virus, and M. gallisepticum were each tested against reference sera with cELISA, ProFLOCK, and Avivac kits. Negative sera from SPF chickens were also included in each test. PI values of each antiserum were expressed as the mean value from duplicate tests by each laboratory. The samples were classified as having positive seroreactivity for PI values greater than 50 (cELISA), 338 (ProFLOCK), or 15 (Avivac).
Seroconversion test after challenge with AIV H9N2 strain
Seroconversion tests following inoculation with the virulent AIV H9N2 strain were performed in chickens. 6 week old SPF chickens were inoculated with 0.2 mL of AIV H9N2 strain (108.1EID50/0.1 mL) via intranasal (0.1 mL/chicken) or oral (0.1 mL/chicken) routes. Each chicken was raised up to 20 days post-inoculation, and 2 mL of blood was collected from wing vein at 1 to 2 day intervals following AIV challenge. The sera were then AIV titer tested by HI and cELISA.
Seroconversion test after vaccination with two different avian influenza viruses subtypes
Seroconversion tests after AIV vaccination were performed by 2 laboratories, GCVP of Korea (H9N2) and ARRIAH of Russia (H5N1). At GCVP, commercial layer hens were vaccinated with an AIV vaccine (GCVP, Korea) prepared with an inactivated H9N2 strain (210 HA unit/dose) and aluminum hydroxide gel. The chickens were inoculated intramuscularly with 0.5 mL of vaccine per chicken, with a boost 2 weeks after the first vaccination with same volume of the vaccine. Sera were taken at before 1 day of prevaccination via wing vein, and at 2 and 4 weeks after the first vaccination with the same method of the first blood collection. All sera were tested for seroconversion by HI and cELISA.
At ARRIAH of Russia, chickens, geese and ducks were inoculated with an inactivated H5N1 vaccine. The H5N1 vaccine was produced from strain A/Duck/Novosibirsk/2/05 (27 HA unit/dose).
Chicken sera were taken from wing vein (2 mL of blood/each sampling) at 10 and 28 days post-vaccination; goose and duck sera were taken at 30 days post-vaccination. Chicken sera were tested with HI, cELISA, Indirect ELISA (ProFLOCK), and Avivac; goose and duck sera were tested with HI and cELISA. The ProFLOCK and Avivac ELISAs were in an indirect format that did not allow us to test goose or duck.
Sensitivity and specificity test
The cELISA was tested with 3,510 sera from diverse species, including chicken (n = 1,782), duck (n = 1031), turkey (n = 213), goose (n = 25), horse (n = 63), quail (n = 46), grey partridge (n = 38), red partridge (n = 5), pheasant (n = 18), swan (n = 4), guinea fowl (n = 19), and swine (n = 266). To determine relative sensitivity and specificity of the cELISA, HI (positive, ≥ 1 : 16), AGP, IZS ELISA, and SRH (in the case of horse sera) were compared to results obtained by the cELISA.
Flock sensitivity
Flock sensitivity was determined from chickens showing clinical signs of low pathogenic avian influenza infection, with chickens chosen from 12 flocks spread over 10 farms. Antibody titers from these sera were analyzed with HI (H9 antigen used), cELISA, and a commercially-available indirect ELISA kit.
Results
Seroreactivity with reference sera
cELISA successfully detected antibodies against the following avian influenza virus strains: H1N1, H2N3, H3N8, H4N8, H5N1, H5N2, H5N3, H5N9, H6N2, H7N1, H7N3, H7N7, H8N4, H9N7, H10N1, H11N6, H11N9, H12N5, H13N6, H14N5, and H15N9. No cross-reactivity was observed with antibodies against NDV, EDS-76 virus, M. gallisepticum, or SPF chicken serum (Table 1). Indirect ELISAs were not applicable to any reference sera other than chicken (data was not shown), due to the requirement of species-specificity conjugates.
Seroconversion after AIV challenge
Seroconversion was detected by cELISA 4 days post-challenge in 2 out of the 5 chickens. The HI test (positive result for titers > 23) detected seroconversion in only 1 of the 5 chickens by day 4. At six days post-challenge, both tests revealed that all chickens had seroconverted to AIV H9N2. When the viral titer was lowered to 104.5EID50/0.1 mL for intranasal inoculation, seroconversion was also detected within 4 days post-challenge (date not shown). The patterns of serum antibody titers with cELISA and HI test are presented in Fig. 1.
Seroconversion test after inactivated AIV vaccine inoculation
All chickens vaccinated with inactivated AIV H9N2 vaccine seroconverted according to the cELISA within 14 days post-vaccination, while all the non-vaccinated birds maintained their seronegative status. This result was in agreement with results obtained by HI test (Table 2).
In the experiment with AIV H5N1 vaccination, all the chickens seroconverted by 10 days post-vaccination according to cELISA, while only 1 of 5 chickens tested seroconverted by 10 days with the HI test. Twentry-eight days post-vaccination, all inoculated chickens had seroconverted according to either the cELISA or HI test. The results from indirect ELISA methods showed significant correlation with the chicken cELISA results (R value > 0.9). The sera from geese and ducks also showed seroconversion compared with pre-immune sera at 30 days post-vaccination when tested by cELISA, but only 4 of the 8 geese, and 2 of the 5 ducks showed seroconversion by HI test (Table 3).
Specificity and sensitivity test in variable species
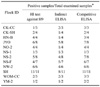
The cELISA was tested with 3,358 sera originating from various species, including chicken, duck, turkey, goose, horse, quail, grey partridge, red partridge, pheasant, swan, guinea fowl, and swine. The cELISA results were compared against those obtained via HI, IZS ELISA, AGP, and SRH. The overall sensitivity and specificity results for these diverse species are presented on Table 4.
Considering the HI as the "gold standard" for seroconversion testing, the sensitivity and specificity for the cELISA as compared with the HI test were 95% and 96% in chicken, 86% and 88% in duck, 97% and 100% in turkey, 100% and 87% in goose, and 91% and 97% in swine, respectively. Both the sensitivity and specificity of the cELISA increased when compared with IZS ELISA in the duck, turkey, goose, and grey partridge sera samples. The results of AGP test against duck and turkey sera also showed significant correlation (R value > 0.9) with the results of cELISA. In the red partridge and swan, the number of sera tested was too low to calculate specificity or sensitivity with any significance. Only positive samples of the horse sera were evaluated, and these were in 100% agreement.
Flock sensitivity
All 12 flocks had a positive reaction against avian influenza virus, as determined by HI titer or ELISA. Specially, the cELISA correlated with the HI test better than the commercially-available indirect ELISAs, with 100% flock sensitivity (Table 5).
Discussion
There are many methods available for detecting avian influenza viral antigens or antibodies in the sera of animals. Antigen-capture ELISAs targeting AIV NP [3], and RT-PCR-based methods have been developed [5,9,16] for virus identification. However, because influenza viruses can clear the bloodstream within one week of clinical signs of infection, antigen detection methods have a short window of efficacy in the large-scale surveillance of herds [3]. Therefore, tests for influenza virus antibodies are preferred for flock surveillance, particularly in large population systems like chickens, swine and other avian livestock [16]. The AGP test is one method used to test for all influenza virus type A-specific antibodies, but this test has several disadvantages, including low sensitivity and a long processing time for test results [10,15]. The competitive ELISA method may be a useful and effective method for the avian influenza surveillance system.
Based on our results monitoring serum antibody titers after viral challenge and vaccination, seroconversion detection time was adequate when using cELISA compared to the HI test. Furthermore, the overall patterns of serum antibody titers demonstrated that competitive ELISA is more sensitive than the HI test. These seroconversion and sensitivity results indicate that the competitive ELISA method could be used to study seroepidemiology of diverse influenza viruses.
The cELISA results for reference sera were in close agreement with those from the HI test, suggesting that that cELISA could detect avian influenza virus antibodies with high fidelity and sensitivity. Optimal sensitivity and specificity of the cELISA compared to the HI test or IZS ELISA were also obtained from the sera of multiple species, including chickens, ducks, geese, guinea fowl, grey partridge and swine. In other avian species (specifically, turkeys, quails and pheasants), the sensitivity of the cELISA was high, but specificity was relatively low. This may indicate that the cELISA should be performed for surveillance, followed by a conventional standard test when specific species need to be tested.
A high degree of sensitivity for AIV detection in positive flocks was also displayed by the cELISA. cELISA detected positive sera with more sensitivity than either the HI test or indirect ELISA. With its high agreement with the HI test in the reference sera and the flock sensitivity test, cELISA could be used to conveniently and quickly screen the avian influenza status of a chicken flock.
Antigenic drift, shift, and re-assortment make it realistic that the avian influenza virus could become infective to a new species. An ideal influenza surveillance system would permit testing of diverse species. The cELISA evaluated in this study showed high detection sensitivity to antibodies from diverse influenza types across various species. This commercially-available cELISA could therefore be used for large-scale surveillance of various species to predict and prevent influenza outbreaks, especially in chickens, ducks and swine.





 XML Download
XML Download