

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Since the production of the first cloned animal from adult somatic cells in sheep, somatic cell nuclear transfer (SCNT) has been used to study cellular processes and embryo physiology, including interactions between somatic cells and the oocyte cytoplasm, and the nuclear remodeling and reprogramming of somatic cells [10,21]. In addition, SCNT has been used in the production of transgenic animals, bio-organ donors, and the multiplication of endangered species and valuable animals [6,17,19]. Recently, it has been shown that bovine, porcine, and rabbit oocytes can support the remodeling and reprogramming of somatic cells from different species, and that these hybrid embryos can develop to the blastocyst stage. Interspecies SCNT (iSCNT) offers a distinct advantage for the production of cloned embryos of exotic or endangered species from which we cannot obtain enough oocytes for research or practical applications. The successful development of iSCNT embryos to the blastocyst stage has been reported in cat [28], gaur [18], human [5], and cattle [29] using rabbit, bovine, or pig oocytes as recipient cytoplasts.
Whales differ from other mammals in terms of their follicle size, serum and follicular fluid compositions, and the oocyte maturation process [14,22]. In contrast to other domestic and experimental animals, few studies of whales have been conducted in the fields of gamete biology and assisted reproductive technology due to difficulties in obtaining somatic cells, spermatozoa, and oocytes. Several studies have attempted to establish a system for the in vitro production (IVP) of whale embryos by the in vitro maturation (IVM) of immature whale oocytes, in vitro fertilization (IVF), or intracytoplasmic sperm injection (ICSI) using Antarctic minke whale spermatozoa [1,8,9,13]. As a result, it is possible to produce whale embryos that develop to the 2- to 16-cell stage by IVP. Notwithstanding the establishment of the whale IVP system, the developmental capacity of whale IVF or ICSI embryos is quite low and no blastocyst formation has been reported following in vitro culture (IVC) of whale embryos. Ikumi et al. [13] first produced cleavage-stage embryos of Antarctic minke whale (Balaenoptera (B.) bonarensis) embryos via intracytoplasmic injection of whale donor cells into bovine and porcine oocytes. Most studies of whale reproductive technology have been performed in Antarctic minke whale; in contrast, there are no reports of the production of sei whale embryos by SCNT. Furthermore, the developmental competence of iSCNT whale embryos produced by subzonal cell injection and electric membrane fusion has not been analyzed.
In the present study, we attempted to produce cloned sei whale (B. borealis) embryos by iSCNT using enucleated pig oocytes as recipient cytoplasts and to improve their developmental capacity by modifying the medium composition and the culture procedure. For this purpose, iSCNT sei whale embryos were produced via the subzonal injection of fetal whale somatic cells into enucleated pig oocytes followed by electric membrane fusion. The effect of osmolarity and macromolecules in the culture medium on the in vitro developmental capacity of the embryos was investigated.
Materials and Methods
The present study was approved by the Animal Experimental Committee of Obihiro University of Agriculture and Veterinary Medicine, Japan, and conducted in accordance with the Guiding Principles for the Care and Use of Research Animals.
Culture media
Unless otherwise stated, all chemicals were purchased from Sigma-Aldrich (USA). The base medium for IVM of the pig oocytes was medium-199 supplemented with 0.1 mM cysteamine, 0.91 mM pyruvate, 3.05 mM glucose, 10 ng/mL epidermal growth factor, 75 µg/mL kanamycin, and 10% (v/v) newborn calf serum (Invitrogen, USA). Porcine zygote medium (PZM)-3 medium [31] containing 2.77 mM myo-inositol, 0.34 mM trisodium citrate, and 10 µM β-mercaptoethanol was used as the IVC medium for embryo development.
Oocyte collection and IVM of pig oocytes
Ovaries were obtained from prepubertal gilts at a local abattoir. The follicular contents were aspirated from superficial follicles (3~8 mm in diameter) in the ovaries with an 18-gauge needle attached to a 6-mL disposable syringe. Cumulus-oocyte complexes (COCs) with more than three layers of compact cumulus cells and a uniform ooplasm were selected and washed three times in HEPES-buffered Tyrode's medium (TLH) containing 0.05% (w/v) polyvinyl alcohol (TLH-PVA) [2] and then washed once in IVM medium. Groups of 70-80 COCs were placed into the wells of a four-well multidish (Nunc, Denmark) containing 500 µL of IVM medium with 10 IU/mL eCG (Teikoku Hormone, Japan) and 10 IU/mL hCG (Teikoku Hormone, Japan). The COCs were then statically cultured at 39℃ in a humidified atmosphere containing 5% CO2. After 22 h of culture, the COCs were washed three times in fresh, hormone-free IVM medium before being cultured again in hormone-free IVM medium for an additional 20 h for SCNT and 22 h for parthenogenetic activation (PA).
Preparation of donor cells
Two sei whales (N = 2) used in this study were captured between May and August 2007 during the second phase of the Japanese Whale Research Program under Special Permit in the west-north Pacific (JARPN-II), which was organized by the Institute of Cetacean Research, Tokyo, Japan. They were killed by explosive harpoons, which have been recognized by the International Whaling Commission as the most humane method. Fibroblasts collected from a male sei whale fetus (16 cm length, 51 g weight) were cultured in Dulbecco's modified Eagle medium (DMEM) containing 1% (v/v) non-essential amino acids solution and 10% (v/v) fetal bovine serum (FBS) for 3 to 6 weeks until a complete monolayer of cells had formed. A suspension of single cells was prepared by trypsinization of the cultured cells, followed by resuspension in TLH containing 0.4% (w/v) bovine serum albumin (TLH-BSA) prior to nuclear transfer. Sei whale fetal fibroblasts used as donor cells were analyzed for their chromosomal ploidy to verify that they had normal diploid (2n = 44) chromosomes. For the chromosomal analysis of donor cells, whale fetal fibroblasts were sub-cultured at passage 4 in DMEM supplemented with 10% FBS. The cells at 60-80% confluence were treated with 0.05 g/mL colcemid for 2~3 h to inhibit the mitotic division. The cells recovered by a trypsin-EDTA solution (Invitrogen, USA) were kept in 0.075 M KCl in distilled water. Then, they were fixed with a methanol-acetic acid (3 : 1) mixture on a glass slide. The slides were conventionally stained with 2% Giemsa (Merck, Germany) in buffered saline (pH 6.8) for 10 min. Sei whale fetal fibroblasts used as donor cells in this study had normal diploid (2n = 44) chromosomes (Fig. 1).
Nuclear transfer
After 42 h of IVM, cumulus cell-free oocytes were incubated for 15 min in manipulation medium (calcium-free TLH-BSA) containing 5 µg/mL cytochalasin B (CB). Following incubation, the oocytes were transferred to a drop of manipulation medium containing 5 µg/mL CB and overlaid with warm mineral oil. The oocytes were enucleated by aspirating the polar body (PB) and a small amount of the adjacent cytoplasm using a 17-µm beveled glass pipette (Humagen, USA). The oocytes were then stained with 5 µg/mL Hoechst 33342 and checked for enucleation under an epifluorescence microscope. Next, single fetal whale fibroblasts were inserted into the perivitelline space of each oocyte. The oocyte-cell couplets were then placed in a 1-mm fusion chamber and overlaid with 1 mL of 280 mM mannitol containing 0.001 mM CaCl2 and 0.05 mM MgCl2 as previously described [30]. Membrane fusion was induced by applying an alternating current field with 2-V cycling at 1 MHz for 2 sec, followed by two or three pulses of 170-175 V/mm direct current (DC) for 15 to 25 µsec using a cell fusion generator (LF101; NepaGene, Japan). The oocytes were then incubated for 1 h in TLH-BSA and evaluated for fusion under a stereomicroscope. Immediately after incubation, the reconstructed oocytes were activated with two DC pulses of 120 V/mm for 60 µsec in 280 mM mannitol containing 0.01 mM CaCl2 and 0.05 mM MgCl2. For PA, oocytes with the first PB at 44 h of IVM were activated using a pulse sequence identical to the one used to activate the SCNT oocytes.
Post-activation treatment and embryo culture
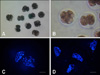
Following electrical activation, the iSCNT whale embryos and PA pig embryos were treated with 5 µg/mL CB in IVC medium for 4 h. Post-activation, the embryos were washed three times in fresh IVC medium, transferred to 30-µL IVC droplets under mineral oil, and then cultured at 39℃ in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2 for 7 days. Embryo cleavage (Figs. 2A and B) and blastocyst formation were evaluated on Days 2 and 7, respectively, with the day of SCNT or PA designated as Day 0. At the end of IVC, the iSCNT embryos were stained with 5 µg/mL Hoechst 33342, the stained nuclei were counted, and their morphology analyzed under an epifluorescence microscope (Figs. 2C and D).
Experimental design
Four experiments were designed to examine the feasibility of iSCNT using pig oocytes for the production of whale embryos and to improve the developmental capacity of iSCNT whale embryos by modifying the culture conditions. The SCNT method used in this study was essentially the same as in our previous SCNT studies [25,26], where SCNT pig embryos were routinely developed to the blastocyst stage. Therefore, only PA embryos were used as a control to verify the stability of the IVC system. In the first experiment, two fusion conditions were compared to determine which would be better for the induction of membrane fusion in whale cell-pig oocyte couplets. Based on our results, two DC pulses of 170 V/mm (25 µsec each) were used in our subsequent experiments. In the second experiment, iSCNT whale embryos and PA pig embryos were cultured in media with two different osmolarities (290 and 360 mOsm) and their developmental capacities were examined. The osmolarity was increased by reducing the volume of water in the medium. Osmolarity of medium was not modified in the later experiments because no beneficial effect of increased osmolarity was found in the second experiment. In the third experiment, iSCNT whale and PA pig embryos were cultured in media containing 0.3% (w/v) BSA, 50% (v/v) FBS, or 50% (v/v) fetal whale serum (FWS) to examine the effect of macromolecule content of the culture medium on embryo development. FWS was prepared from blood collected from the umbilical cord of a male sei whale fetus (231.5 cm in length). The osmolarity of FWS used in this study was 326.2 ± 0.4 mOsm. The blood was allowed to clot by standing at room temperature, after which the serum was recovered by centrifugation at 500 × g for 10 min; the FWS was then stored at -20℃ until use. In the fourth experiment, a two-step culture method was applied to test whether exposure of the embryos to FBS or FWS from Day 3 of culture would stimulate the development of iSCNT whale embryos beyond the developmental arrest at the 4-cell stage. In this experiment, the concentration of FBS and FWS in culture medium was decreased to 15% (v/v) because 50% FBS and FWS did not show any stimulatory effect on iSCNT embryo development in the preceding experiment.
Results
Effect of the electric conditions on oocyte-cell fusion and the development of iSCNT whale embryos (Experiment 1)
The proportions of survived and fused oocytes were not significantly different between the two electric conditions tested (Table 1). When iSCNT embryos produced by two DC pulses of 170 V/mm for 25 µsec and three DC pulses of 175 V/mm for 15 µsec were cultured, 60 and 65% of the embryos cleaved, respectively, but none of the iSCNT whale embryos developed past the four-cell stage. In comparison, the PA pig embryos produced greater (p < 0.05) cleavage (94%) and blastocyst formation (82%) than the iSCNT whale embryos (Table 2).
Effect of medium osmolarity on the in vitro development of iSCNT whale and PA pig embryos (Experiment 2)
Embryo cleavage and blastocyst formation in the iSCNT whale embryos were not altered by increasing the osmolarity of the medium to 360 mOsm. At 360 mOsm, there was a general increase in the number of embryos that developed past the two-cell stage (p = 0.0781). Among the PA pig embryos, the cleavage rate was not influenced by the osmolarity of the medium, but the blastocyst formation rate (80%) was significantly higher (p < 0.05) at 290 mOsm than at 360 mOsm (43.9%; Table 3).
Effect of macromolecules in the culture medium on in vitro development of iSCNT whale and PA pig embryos (Experiment 3)
As shown in Table 4, iSCNT embryo development was not influenced by the macromolecule content of the culture medium. Supplementation of IVC medium with BSA, FBS and FWS showed similar rates of iSCNT embryo cleavage (81.1%, 78.4% and 70.3%, respectively). In the PA pig oocytes, cleavage was significantly reduced when the oocytes were cultured in the presence of FWS. In comparison, no blastocysts developed in the presence of FBS or FWS. When the nuclear morphology of iSCNT embryos arrested at the two- to four-cell stage on Day 7 of IVC were examined, most of the embryos exhibited nuclei with metaphase-like structures (Figs. 2C and D). Irrespective of the type of macromolecule added to the culture medium, all of the embryos arrested at the cleavage stage showed a similar metaphase-like nuclear morphology.
Effect of a two-step culture method using different macromolecules in the medium on the in vitro development of iSCNT whale and PA pig embryos (Experiment 4)
The culture medium was supplemented with FBS or FWS after two days of culture to determine if it would stimulate the development of iSCNT whale embryos. As shown in Table 5, no stimulatory effect was found on embryo development by serum supplementation. Even in the PA pig oocytes, no blastocyst formation was observed in the presence of FBS or FWS.
Discussion
We found that porcine IVM oocytes could support in vitro development of iSCNT sei whale embryos to the four-cell stage, which indicates porcine oocytes could induce the nuclear remodeling and reprogramming of sei whale somatic cells. In addition, it was found that most of the cleaved whale embryos that arrested at the two- to four-cell stage showed metaphase-like nuclear structures, which may suggest an in vitro developmental block. To the best of our knowledge, this is the first study to report the successful production of iSCNT sei whale embryos via the subzonal injection of donor cells followed by electrofusion.
Whale-porcine iSCNT embryos were produced by the subzonal injection of whale donor cells followed by electric membrane fusion. The rate of fusion between the whale somatic cell and pig oocyte membranes was similar to that between pig donor cells and pig oocytes. The electric field strengths (170~175 V/mm) and pulse durations (15~25 µsec) used in this study were similar to those used previously for pig SCNT [26], and have been shown to be acceptable for the membrane fusion of whale somatic cells with pig oocytes. In iSCNT embryo culture, it is common to use a medium specific for the oocytes used as recipient cytoplasts [13,29]. Therefore, in the present study, iSCNT sei whale embryos were cultured in PZM-3 medium, which was developed for the culture of pig embryos [31]. When the electrically fused embryos were cultured, 60-81% of the iSCNT whale embryos developed to the two- to four-cell stage but no blastocyst formation was observed. In contrast, the PA pig embryos showed high rates of cleavage (94~100%) and blastocyst formation (64.7~82%). It is unclear whether the developmental arrest at an early stage of cleavage was due to incompatibility between the whale somatic cells and pig oocytes or to suboptimal culture conditions for the hybrid embryos.
To improve the low developmental competence of the iSCNT whale embryos, we made several modifications to the medium, including varying the osmolarity and macromolecule content. We previously showed [22] that the osmolarities of follicular fluid (363.3~388.9 mOsm) and umbilical serum (379.5 mOsm) in Antarctic minke whales exceeded the serum or plasma value in other domestic species (300 mOsm) [11,23]. Based on the high osmolarity of whale follicular fluid and serum, we designed a new medium with a high osmolarity for the culture of iSCNT whale embryos. More of the embryos that were cultured at 360 mOsm developed past the two-cell stage, but none of the embryos developed beyond the four-cell stage, even though the osmolarity of the culture medium was similar to whale follicular fluid or umbilical serum. In contrast, the development of the pig PA oocytes was significantly influenced by the osmolarity of the culture medium. In this study, osmolarity was increased by reducing the volume of water in the medium which might increase the concentration of medium components and then affect embryonic development. It has been shown that fetal serum is rich in embryotrophic substances, including growth-promoting factors and amino acids, and that it stimulates embryonic development in many species [4,12,20,24]. In the present study, there was no beneficial effect of the addition of FWS or FBS to the culture medium on the development of iSCNT whale embryos. Unexpectedly, FBS supplementation was unable to support PA pig embryo development to the blastocyst stage although the same batch of FBS supported blastocyst formation of bovine IVF or PA embryos in our preliminary experiment (unpublished data).
It has been well known that FBS has biphasic effects on the bovine and porcine embryo development depending on the time of supplementation. FBS is supplemented to the culture medium routinely at the later stage of preimplantation development in pigs because exposure of early porcine embryos is detrimental to blastocyst formation in vitro [7,16]. In addition, the effect of FBS varies depending on different batches [15,27]. In this study, pig PA embryos were cultured in a medium containing FBS from the one-cell stage. Therefore, the early exposure of PA embryos to FBS or undesirable batch of FBS might inhibit blastocyst formation. Due to the limited amount of information available concerning the optimal medium osmolarity and the effect of fetal serum on whale embryo development, we are currently unable to explain why no embryotrophic effect of osmolarity and macromolecule composition was observed.
A fairly large proportion of the iSCNT whale embryos were able to develop to the cleavage stage (60~81%), similar to the percentage observed following the culture of in vitro-fertilized or SCNT pig embryos [3,32]. Although it was not possible to compare them directly, the cleavage rate of the iSCNT sei whale embryos in this study was higher than that (25.0~42.7%) in a previous study of iSCNT Antarctic minke whale embryos created by the direct injection of somatic cells into the cytoplasm of enucleated porcine oocytes [13]. This suggests that the subzonal injection of donor cells with electrofusion may be more efficient for the production of large numbers of iSCNT whale embryos. Despite the relatively normal morphology of the iSCNT whale embryos, all of the cleaved embryos arrested at the two- to four-cell stage possessed nuclei with metaphase-like structures. Therefore, we assumed that the developmental block occurred at or just before metaphase during the second or third cell cycle. Accordingly, a two- step culture method was applied, in which two-cell iSCNT embryos that had completed their first cell cycle were cultured in fresh medium containing FBS or FWS, and their progress was monitored through the second cell cycle. Although the two-cell embryos were exposed to FBS or FWS in the culture medium before their progression into the next cell cycle, it was not possible to overcome the developmental arrest. Despite our efforts in this study, the developmental competence of the whale-porcine hybrid embryos was not greatly affected by modification of the whale embryo culture system. It is unclear whether the low developmental ability of the embryos was due to an intrinsic problem such as the incompatibility between whale somatic cells and pig oocytes or a suboptimal culture environment. Additional studies are needed to overcome the developmental arrest at the two- to four-cell stage in hybrid whale embryos by utilizing more compatible oocytes than pig oocytes as recipient cytoplasm for iSCNT.
In summary, we produced iSCNT whale embryos by subzonal injection and electrofusion. The pig oocytes used in this study supported the remodeling and reprogramming of the whale somatic cell nuclei, but they could not fully support the development of the embryos to the blastocyst stage. The SCNT technique developed in this study using electrofusion after subzonal cell injection may be used for the mass production of cloned whale embryos in the near future.






 XML Download
XML Download