

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Apoptosis, also called programmed cell death, is essential for the homeostasis of normal tissues. Altering the level of apoptosis is involved in various diseases including cancer, viral infections, autoimmune diseases, neurodegenerative disorders and AIDS [22]. Therefore, controlling the apoptotic process may provide a critical leverage point for the treatment of various diseases.
Akt, also named protein kinase B, is known to be a critical target for cancer intervention. It is activated downstream of phosphoinositide 3-kinase (PI3K) by phosphorylation on two regulatory residues, Thr-308 and Ser-473 [3]. The activation of Akt plays a critical role in fundamental cellular functions such as cell proliferation and survival by phosphorylating a variety of substrates. Constitutively active Akt results in augemented resistance against apoptotic cellular insults, such as growth factor deprivation, UV irradiation or loss of matrix attachment [15]. Akt activation is found in many types of human tumors including breast cancer, lung cancer, melanoma and leukemia [7,16].
Extracellular signal-regulated kinase (ERK)1/2 is also crucial molecule in cell proliferation and carcinogenesis. It is activated by dual phosphorylation on both Thr202 and Tyr204 residues. Activated ERK1/2 has been reported in a variety of human tumor cell lines [8] and epithelial cancer tissues such as breast [1], kidney [17], colon [20], head and neck [2] and small-and non-small-cell lung cancer [4]. In many cases, ERK activation protects cells from drug-induced cell death [21]. A number of studies have indicated that the phosphorylation of ERK promotes cell survival by inhibiting apoptosis under various pathological conditions [5].
Tetrandrine (TET), a bis-benzylisoquinoline alkaloid from the root of Stephania tetrandra, has been used in China for several decades for the treatment of arthritis, arrhythmia, inflammation and silicosis [18]. TET was also reported to inhibit cellular proliferation in various cancer cell types [14]. However, the precise mechanisms by which TET inhibits tumor cell growth remain to be elucidated. In this study, therefore, we investigated the effects of tetrandrine on PI3K/Akt and ERK pathways in A549 human lung carcinoma cells. Here, we report that TET-induced apoptosis is closely associated with Akt-ERK crosstalk.
Materials and Methods
Reagents
TET was purchased from Sigma-Aldrich (USA). Anti-Bid, anti-Bax, anti-Bcl-xL, anti-Akt, anti-phospho-Akt Thr-308, Ser-473, anti-ERK and anti-phospho-ERK antibodies for Western blot analysis were purchased from Santa Cruz Biotechnology (USA). All reagents used in this study were reagent grade or better.
Cell culture and treatment
A549 human lung carcinoma cells were obtained from American Type Culture Collection (USA) and grown in RPMI-1640 medium supplemented with 10% fetal bovine serum (FBS; Hyclone Lab, USA). Cells were incubated at 37℃ in a humidified atmosphere with 5% CO2. For TET treatment, cells were plated at a density of 2 × 106 cells per T-75 cm2 culture flask, stabilized for 24 h and then treated with TET for the times and concentrations indicated. TET was dissolved in DMSO (Sigma-Aldrich, USA) at 20 mM as a stock solution and diluted for further analysis.
The concentration-dependent effect of TET on the inhibition of A549 cell proliferation
The impact of TET on the viability and proliferation of A549 cells was determined using the (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT) assay. Briefly, cells were plated in 96-well culture plates (5 × 104 cells/well). After 24 h incubation, the cells were treated with TET (0, 5, 10, 20, 30, 40, 50 or 60 µM) for the indicated times. After treatment, 10 µl of MTT solution (1 mg/ml in PBS) were added to each well and the plate was incubated for 4 h at 37℃. To achieve solubilization of the formazan crystal formed in viable cells, 100 µl of DMSO were added to each well. The plate was shaken for 15 min at room temperature and the absorbance was measured using a microplate reader (Bio-Rad, USA) at a wavelength of 595 nm.
Western blot analysis
Protein concentration was determined using a Bradford analysis kit (Bio-Rad, USA). Equal amounts of protein were separated on a 12% SDS polyacrylamide gel and transferred to a nitrocellulose membrane (Hybond ECL; Amersham Pharmacia, USA). The blots were blocked for 2 h at room temperature with blocking buffer (10% nonfat milk in TTBS buffer containing 0.1% Tween 20). The membrane was incubated at room temperature for 1 h with specific antibodies. The antibodies were used at 1 : 1,000 dilutions as specified by the manufacturer. After washing with TTBS, the membrane was incubated with a horseradish peroxidase-labeled secondary antibody and visualized using the Westzol enhanced chemiluminescence detection kit (Intron, Korea). The bands were detected with LAS-3000 (Fujifilm, Japan).
Flow cytometric detection of apoptosis
The percentage of apoptotic cells was determined by staining cells with annexin V-FITC and propidium iodide (PI). The annexin V-FITC apoptosis detection kit was purchased from Calbiochem (Canada). After incubation, cells were transferred to a microfuge tube, washed with ice-cold PBS, then resuspended in 0.5 ml cold × 1 binding buffer, followed by the addition of 1.25 µl of annexin V-FITC. The mixture was incubated at room temperature for 15 min in the dark. After adding PI, the samples were analyzed by FACS Calibur Flowcytometry (Becton Dickinson, USA).
Selective inhibitor study
The mitogen-activated protein kinase kinase (MEK1/2) inhibitor PD98059 and PI3K inhibitor LY294002 were purchased from Tocris (USA) and Calbiochem (Germany), respectively. Stock solutions were prepared in DMSO. The highest concentration of DMSO used was 0.2%. For the co-treatment experiments using TET and an inhibitor, cells were preincubated with either PD98059 (50 µM) or LY294002 (20 µM) for 1 h prior to TET treatment.
Statistical analysis
Result are shown as the mean ± SE. Statistical analyses were performed following ANOVA (MS-Excel 2003; Microsoft, USA) for multiple comparisons or Student's t-test when the data consisted of only two groups. The differences between groups were considered significant at p < 0.05 and p < 0.01 as indicated.
Results
To determine the effects of TET on cell viability, the MTT assay was performed on A549 cells treated with various concentrations of TET. The cells were exposed to 0-60 µM of TET for 24 h and 48 h. TET treatment significantly reduced the rate of cell proliferation compared to that of control cells in both time-/concentration-dependent manners. The reduction of cell proliferation and thus cell viability following treatment with 30 µM TET was roughly 59% at 24 h (Fig. 1A) and 43% at 48 h (Fig. 1B). These results led us to use 30 µM of TET for further studies.
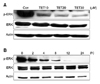
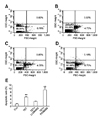
Since the MTT assay is a measure of total cell numbers and the results reflect changes in both cell proliferation as well as apoptosis, we next characterized the specific effects of TET on levels of apoptosis. To do this, a flowcytometric detection method was used after cells were treated with 30 µM TET for 12 h and 24 h. The lower right quadrant (Annexin V positive and PI negative) represents the percentage of apoptotic cells with preserved plasma membrane integrity whereas the upper right quadrant (Annexin V positive and PI positive) refers to necrotic or apoptotic cells with a loss of plasma membrane integrity. It was determined that most cells were alive since untreated cells were not stained with Annexin V or PI (Figs. 2A-C). The apoptotic fraction of cells treated with TET is represented in Fig. 2D. Treatment with TET caused apoptosis in a time-dependent manner; approximately 0.42 ± 0.07 (0 h), 1.86 ± 0.11 (12 h) and 4.88 ± 0.95% (24 h) of apoptotic cells were observed (Fig. 2D). Also, treatment with TET significantly decreased the expression level of the anti-apoptotic protein Bcl-xL in a concentration-dependent manner whereas the levels of the pro-apoptotic protein Bax remained unchanged (Fig. 3A). These tetradine-mediated effects on the apoptosis of A549 cells were clearly observed in time-course study. Treatment with 30 µM of TET resulted in a significant increase in the levels of the pro-apoptotic proteins Bid and Bax whereas the expression levels of the anti-apoptotic protein Bcl-xL decreased in a time-dependent manner (Fig. 3B).
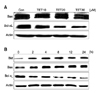
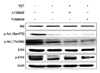
Since Akt is a crucial mediator of carcinogenesis and the phosphorylation of Akt is essential for its full activity and is involved in apoptosis [9], we have measured the potential effects of TET on Akt phosphorylation. TET treatment suppressed Akt phosphorylation at both Thr308 and Ser473 in both time- and concentration-dependent manners, while the total Akt levels remained unchanged (Fig. 4). ERK is also known to be a pivotal factor in carcinogenesis and is closely associated with Akt signaling [19] and therefore the potential effects of TET treatment on ERK signaling were measured. Interestingly, TET also suppressed ERK phosphorylation in both time-/concentration- dependent manners similar to Akt phosphorylation (Fig. 5).
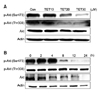
To characterize the relative roles of Akt and ERK on TET-induced apoptosis, two different selective inhibitors (LY294002 for PI3K pathway, PD98059 for MEK/ERK pathway) were used. TET alone increased apoptosis when compared to control (Figs. 6A, B and E). However, the fraction of apoptotic cells in samples co-treated with TET and the ERK inhibitor PD98059 was significantly increased compared to treatment with TET alone (Figs. 6D and E). Interestingly, cells co-treated with TET and the PI3K inhibitor did not manifest such synergetic effects (Figs. 6C and E). Our results strongly suggest that the inactivation of ERK may play an important role in TET-induced apoptosis. TET alone was enough to suppress the phosphorylation of Akt at both Ser473 and Thr 308 (Fig. 7) in both time-course as well as dose-response studies (Fig. 4). The expression of phosphorylated Akt was further suppressed by co-treatment with TET and LY294002 or PD98059 (Fig. 7). Very similar phenomena were found in terms of ERK phosphorylation (Fig. 7).
Discussion
Lung cancer is a major cause of cancer-related mortality worldwide. Lung cancer has proven difficult to control with conventional therapeutic and surgical approaches, and the prognosis is poor with an overall 5 year survival rate of 10-14% in the USA [11]. Therefore, it is clear that novel and more effective treatments are needed to improve the outcome of therapy. In this respect, the use of naturally occurring or synthetic agents to prevent, inhibit or reverse lung carcinogenesis would greatly benefit public health. TET is a promising phytochemical agent that has recently attracted interest because of its cancer chemopreventive potential. In this study, TET, a candidate for use as a lung cancer chemopreventive agent, was characterized in the cell line A549.
Growing evidence has demonstrated that PI3K/Akt pathways are involved in several types of carcinogenesis. The activation of Akt causes malignant transformation in in vitro and in vivo mouse models of various human cancers [10]. In our study, TET suppressed Akt phosphorylation at Ser473 and Thr308 and inhibited lung tumorigenesis. The anti-tumor activity of TET appears to be mediated by the suppression of Akt phosphorylation because Akt requires phosphorylation of both Thr308 and Ser473 for full activity [24]. Our finding is clearly supported by previous reports that Akt activation is an early event in lung tumorigenesis [6], and that blocking Akt activity could suppress the progression of lung adenocarcinoma [12]. TET, therefore, may be an excellent lung cancer chemopreventive agent because one of the most promising molecules for chemoprevention and for the treatment of lung cancer targeting Akt.
Akt and ERK are both important signaling molecules that promote survival in different types of cancer. Spatiotemporal control of the ERK signal pathway is a key factor for determining the specificity of cellular responses including cell proliferation, cell differentiation and cell survival. The fidelity of this signaling is tightly regulated by docking interactions as well as scaffolding. The subcellular localization of ERK is controlled by cytoplasmic ERK anchoring proteins that have a nuclear export signal such as MEK. In quiescent cells, ERK localizes to the cytoplasm. In response to stimulation, activated ERK translocates to the nucleus [23]. To get detailed information about the relative roles of such signaling in lung cancer cell survival, the effect of treatment with TET and the PI3K inhibitor LY29294002 as well as the ERK inhibitor PD 98059 on the expression patterns of Akt within A549 cells was examined. TET treatment induced apoptosis and resulted in a decrease in Akt and ERK expression. PI3K inhibition had no clear synergistic effect on tetradrine-induced apoptosis, however, ERK inhibition resulted in a significant synergistic effect on apoptosis such that the degree of apoptosis was much higher than TET treatment alone and TET with PI3K inhibitor pretreatment. Western blot analysis of Akt and ERK protein levels and activation states confirmed that TET-induced apoptosis may occur under the dual action of ERK and Akt. Taken together, our results suggest that TET induces apoptosis and promotes the down-regulation of Akt expression in A549 lung cancer cells with a close relationship to ERK activity. Our results are further confirmed by other lines of evidence, which indicate that ERK regulates cell death in many cell lines. Increased levels and/ or the activation of ERK have been observed in a number of human cancer cell lines [8].
The evidence presented here suggests that TET deactivates Akt and synergistically promotes apoptosis through the inhibition of ERK. Such selective down-regulation of Akt activity and facilitating apoptosis indicates the potential utility of TET as a promising target for the prevention of lung cancer because Akt is likely to be an important factor in the early progression of lung carcinoma. The data presented provide evidence that TET selectively inhibits the proliferation of lung cancer cells by blocking Akt activation and that it facilitates apoptosis by ERK inhibition. Because Akt activity alters the sensitivity of non-small cell lung cancer cells to chemotherapeutic agents and irradiation [13], lung cancer treatment with TET may enhance the efficacy of chemotherapy and radiotherapy, and increase the apoptotic potential of lung cancer cells.


 XML Download
XML Download