

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Galectin-3 is a β-galactoside-binding animal lectin that contains carbohydrate-recognition domains and displays multiple related functions [2]. Extracellular galectin-3 mediates inflammation [1], whereas intracellular galectin-3 regulates cell growth and anti-apoptosis, and modulates cell adhesion, thus inducing cell migration [16]. Galectin-3 has been found in the cytoplasm of various cell types, including inflammatory cells such as macrophages, dendritic cells, mast cells, neutrophils, and eosinophils [14,15]. The expression of galectin-3 is known to be upregulated following certain bacterial [5] and parasitic infections [24]. In addition, it is accumulated in phagosomes containing mycobacterium during the course of an infection [3]. Here, we propose that galectin-3 is associated with the formation of granulomatous inflammatory lesions in chronic diseases including paratuberculosis.
Paratuberculosis, or Johne's disease, is a chronic, granulomatous enteritis found in wild and domestic ruminants. The causative agent is Mycobacterium avium subsp. paratuberculosis [18], which is a slow-growing facultative intracellular bacterium that persists within macrophages in the intestinal tract for several years before clinical onset [6]. The symptoms of clinical paratuberculosis are chronic diarrhea and progressive weight loss; subclinically infected animals may display decreased production [22]. The gross lesions are characterized by a segmental thickening of the intestine and mesenteric lymphadenopathy [18].
The aim of this study was to examine the immunohistochemical localization of galectin-3 in granulomatous intestines of bovines infected with paratuberculosis to determine whether galectin-3 was involved in the formation of granulation tissue in chronic disease. rpoB, encoding the ß subunit of RNA polymerase [4], PCR-plasmid TA cloning-sequencing for Mycobacterium species was used to diagnose the presence of paratuberculosis [27].
Materials and Methods
Tissue samples (n = 3) from the small intestines, mainly ileum, and mesenteric lymph nodes were obtained from cows at slaughter that were suspected to be infected with paratuberculosis. Tissue samples were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) and processed for paraffin-embedding. Sections (5 µm thick) were cut in a Microtome (Leica, Germany), and were routinely examined with hematoxylin and eosin staining and with Ziehl-Neelsen staining for acid-fast bacilli [17]. As a control, bovine ileum without acid-fast bacilli staining was used.
For immunohistochemistry, 5-µm-thick sections of paraffin-embedded bovine ileum and mesenteric lymph nodes were deparaffinized using xylene and pure ethanol before exposed to citrate buffer (0.01 M, pH 6.0). Samples were then heated in a microwave oven for 3 min. All subsequent steps were performed at room temperature. The sections were treated with 0.3% hydrogen peroxide in methyl alcohol for 20 min to block endogenous peroxidase activity. After three washes in PBS, the sections were blocked with 10% normal goat serum (Vector ABC Elite Kit; Vector Laboratories, USA), diluted in PBS for 1 h, and then incubated with rat anti-galectin-3 antibody (1 : 5,000) for 1 h. The rat anti-galectin-3 monoclonal antibody (1 mg/mL) was purified from the supernatant of hybridoma cells (TIB-166; ATCC, USA). After three washes in PBS, the sections were incubated with biotinylated goat anti-rat IgG (1 : 100; Vector Laboratories, USA) for 45 min. After three washes in PBS, the sections were incubated with the avidin-biotin peroxidase complex (Vector Laboratories, USA), prepared according to the manufacturer's instructions, for 45 min. The peroxidase reaction was developed using a peroxidase substrate kit (Vector Laboratories, USA) according to the manufacturer's protocol. After the completion of color development, the sections were counterstained with hematoxylin (Sigma, USA) for 5 sec, washed in running tap water for 20 min, dehydrated through a graded ethanol series, and then cleared with xylene and mounted with Canada balsam (Sigma, USA). As a negative control, the primary antibody was omitted.
For the diagnosis of paratuberculosis from paraffin embedded tissues, the paraffin-embedded specimens were deparaffinized with xylene and pure ethanol. DNA was extracted using the previously described bead beaterphenol extraction method [10]. The DNA pellet obtained was used as a template for PCR. The rpoB PCR was carried out as described previously [10]. The PCR products obtained were electrophoresed in a 1.5% agarose gel and purified using a QIAEX II gel extraction kit (Qiagen, Germany). The purified PCR product (5-10 ng) was cloned using a TA cloning kit (Invitrogen, USA) according to the manufacturer's instructions. Three to 10 colonies of transformed Escherichia coli were picked in each reaction, cultured, and used to prepare plasmid DNA with a High Pure Plasmid Isolation Kit (Roche, Germany). The nucleotide sequences of the cloned rpoB DNAs were directly determined from the purified plasmid using M13 forward and reverse primers, which were supplied in the TA cloning kit, a 373A automatic sequencer, and a BigDye Terminator Cycle Sequencing kit (PE Applied Biosystems, UK). For the sequencing reaction, 60 ng of PCR-amplified DNA, 3.2 pmol of either the forward or the reverse primer, and 4 µL of BigDye Terminator RR mix (PE Applied Biosystems, UK) were mixed and adjusted to a final volume of 20 µL by adding distilled water. The reaction was run with 5% (vol/vol) dimethyl sulfoxide for 30 cycles of 15 sec at 95℃, 10 sec at 50℃, and 4 min at 60℃. Both strands were sequenced as a cross-check. The sequences determined (306 bp) were aligned and compared to sequences in GenBank by using the multiple-alignment algorithms in the MegAlign package (Windows version 3.12e; DNASTAR, USA).
Results
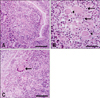
All three cases showed typical granulation tissues in the intestines with a varying degree of inflammation. The histological findings in three cases were similar. Briefly, the lamina propria and submucosa of the small intestine was thickened due to the infiltration of inflammatory cells (Fig. 1A). The thickening of the mucosa was attributable to the accumulation of typical lymphocytes with condensed nuclei and macrophages with foamy, pale cytoplasm (Fig. 1B). Giant cells including more than two nuclei were occasionally found (Fig. 1B, arrowheads). An accumulation of multinucleated giant cells were also found in the mesenteric lymph nodes (Fig. 1C).
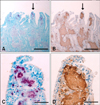
In the acid-fast stained tissue sections of the small intestine, the presence of acid-fast bacilli was confirmed due to its red coloration (Fig. 2A). Acid-fast bacilli were found in various cell types, including macrophages, in the lamina propria and submucosa of the small intestine. The lamina propria just below the epithelium of the villi was intensely stained red, suggesting that acid-fast bacilli were compacted in macrophages within this area (Fig. 2C). In the adjacent tissues of the acid-fast stained section, galectin-3 was strongly expressed in the round cells (typical of macrophages) in the lamina propria and submucosa of the intestines (Fig. 2B) and moderately expressed in the intestinal epithelium. The intense staining patterns of galectin-3 in the villous lamina propria (Fig. 2D) largely overlapped with the acid-fast staining (Fig. 2C).
The rpoB DNA (368 bp) was amplified from the small intestine sample, including ileum. However, due to weak amplification, PCR-direct sequencing was not possible. Therefore, the PCR products obtained were cloned into a TA plasmid for sequencing, and the inserted DNA sequences of the sample were compared with the sequences of 44 reference strains of mycobacteria and sequences in the GenBank database. The sequence showed 100% homology with Mycobacterium avium subsp. paratuberculosis (GenBank accession no. AE016958.1). Therefore, the cows were confirmed to be positive for Mycobacterium avium subsp. paratuberculosis.
Discussion
This study confirms for the first time the accumulation of galectin-3-positive macrophages in bovine paratuberculosis-infected granulation tissues, particularly the ileum. Galectin-3 has long been known as an important mediator of macrophage activation [1,7] and postulated to play a critical role in phagocytosis by macrophages [23]. However, the precise mechanisms of how galectin-3 interrupts bacilli digestion by phagosomes within macrophages during paratuberculosis infection remains unknown and requires further study. In addition, galectin-3 has been known to be a major adhesion molecule [8] and involved in the trafficking of inflammatory leukocytes, including activated macrophages [1]. Based on these previous studies, we hypothesize that macrophages continuously accumulate in the granulation tissues.
Following the accumulation of macrophages in the granulation tissues of paratuberculosis, macrophages were found to be alive in the lesions even though they contained bacilli [6]. Bacilli-containing macrophages are thought to survive due to the anti-apoptotic characteristics of galectin-3 [16,21,25]. The molecular mechanism underlying this effect is cytochrome c release from the mitochondria [26], which possibly blocks cell death. In contrast to the increase of galectin-3 in macrophages during paratuberculosis, galectin-3 decreases in the intestinal epithelia of animals with intestinal bowel disease [19], suggesting that the expression of galectin-3 is cell type-dependent.
In addition to the immunological role of galectin-3 in inflammatory cells, galectin-3 has been found in the epithelia of the digestive and respiratory tracts [9,12,20] as well as the reproductive organs [11,13], where mucus is abundant. In the present study, we found galectin-3 localized in the intestinal epithelia, particularly colocalized with mucin. Since galectin-3 is one of the ligands for mucin [8], we postulate that galectin-3 is involved in the mucin-mediated protective mechanisms of epithelial cells.
In conclusion, we postulate that intracellular galectin-3 is involved in the protection of macrophages that contain bacilli, while extracellular galectin-3 is involved in the facilitation of inflammatory cell accumulation. Both machineries synergistically lead to chronic granulomatous bowel disease in bovine paratuberculosis.
 XML Download
XML Download