

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Introduction
Porcine reproductive and respiratory syndrome virus (PRRSV) is a small, enveloped RNA virus [4,39] that is classified as a member of the genus Arterivirus of family Arteriviridae in the order Nidovirales [6]. PRRSV was first isolated almost simultaneously in Europe [40] and the USA [9].
The PRRSV genome is a polyadenylated, single-stranded, non-segmented, positive-sense RNA approximately 15 kb in size and consisting of at least 9 open reading frames (ORFs) [10,27,30,35,41]. ORF1a and 1b are located immediately downstream of the 5' untranslated region (UTR) and involved in virus transcription and replication. ORFs 2 to 7 are located at the 3' end of the genome and are thought to encode for the PRRSV structural protein. Among them, ORF5 encodes an unglycosylated membrane of 17 kD or a glycosylated protein of 25 kD [22,26]. Furthermore, the ORF5 of PRRSV is in an immunologically crucial region which is mainly related with neutralizing antibody formation. In the case of type II PRRSV, ORF5 has been known to contain three B-cell epitopes and two T-cell epitopes [11,31,32,38].
Remarkable genetic differences have been reported among PRRSV. The major two genotypes are European (Type I, Lelystad as prototype) and North American (Type II, VR-2332 as prototype). The similarity between these two genotypes among the ORFs were shown to be 57~59% (ORF7), 70~81% (ORF6), 51~59% (ORF5), 68% (ORF4), 58% (ORF3) and 63% (ORF2) [3,14,23,24,25,28]. Even within the same genotype, high genetic variation among field viruses has been reported all over the world. Type II PRRSV was first isolated in Korea in 1994 [21], with the molecular characterization of Korean PRRSV from 1996 to 2006 based on ORF5 and ORF7 [7,8,43]. These studies suggested that the Korean PRRSVs belonged to the Type II PRRSV and had geographically evolved with their own genetic clusters. However, one group also reported that type I PRRSV also existed in Korea [18].
To control PRRSV, the modified "test and removal method" was successful in the seed stock breeding farm and a supplying boar studs in Korea [42]. Since this method is laborious and time-consuming, most swine farms with continuous flow systems have applied a modified live vaccine (MLV) .
With the widespread use of MLV, the actual efficacy of the vaccine came into question as the sequence analysis based on the ORF5 sequences of Korean PRRSV showed that the genetic divergence was ranged from 1.3 to 12.9% compared to the MLV [7]. Although MLV had an effect to reduce clinical signs and enhance weight gain in the PRRSV-infected farms, the virus shedding of challenged heterologous viruses was not prevented [5].
With these concerns, this study focused on analyzing the patterns of viremia detected post-vaccination, which could represent the active replication of PRRSV, in the either MLV vaccinated or non-vaccinated farms. Furthermore, the full ORF5 sequences acquired from viremic pigs were compared with the reported [7,21] or previously collected data in the laboratory including type I and II PRRSV from 1997 to 2008.
Materials and Methods
Study design
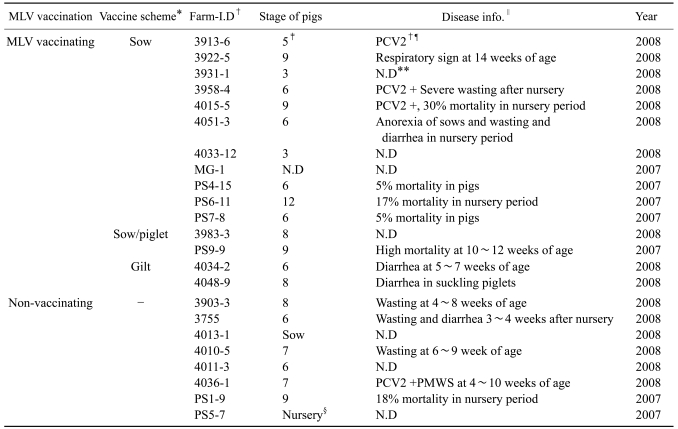
Blood samples were collected from commercial swine farms with information about MLV vaccinating and disease status. In the case of MLV vaccinating farms, blood samples were collected at least 6 months after the first vaccination. If the main clinical signs and PCV2-specific PCR results were available, they were also documented (Table 1).
At first, pilot study was performed to preview the viremic patterns in the MLV vaccinating and non-vaccinating farms. The vaccine schedules used in the MLV-vaccinating farms were one of the sow-only or sow/piglet vaccinations. Among 25 farms, the number of farms with viremic pigs at 3, 6, 9 and 12 weeks of age, including sows, was investigated using a reverse transcriptase nested polymerase chain reaction (RT-nested PCR) method. Serum antibody titer was also evaluated with a commercial enzyme-linked immunosorbent assay (ELISA) kit (Herdchek 2XR, PRRS; IDEXX, USA). The serum IgG titers were expressed as S/P ratio: (Sample O.D - Negative control O.D) / (Positive control O.D - Negative control O.D).
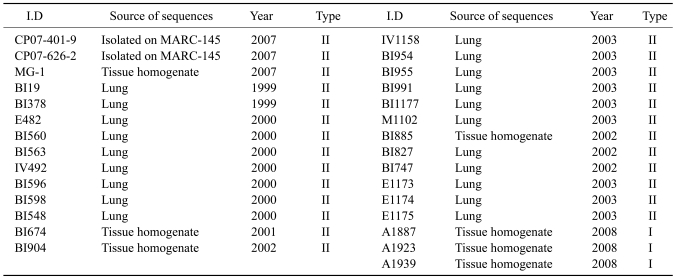
After the pilot study, to know the genetic properties of PRRSV in the viremic sera between MLV-vaccinating or non-vaccinating farms, the viremic samples collected from 2007 to 2008 were further sequenced based on ORF5. A total of 23 PRRSV ORF5 sequences in the viremic sera were obtained. The sequenced data was analyzed with reference and previous laboratory sequence data including type I and II PRRSV from 1997 to 2008, to observe the phylogenetic relationship. The detailed sequence information was presented in Tables 1 and 2.
Except for previously-published reference sequences, the sequence data of type I PRRSV and the other type II PRRSV (not collected from viremic sera) used in this study were collected from 1999 to 2008 in the virus-infected tissues of commercial pigs in Korea. The ORF5 sequences used in this study are available in GenBank accession numbers, from FJ972714 to FJ972766.
Virus isolation
Two Korean field isolates (CP07-401-9 and CP07-626-2, both in 2007) were adapted on MARC-145 cell which was known to be permissive to the PRRSV, especially type II PPRSV [15] and used for genetic analysis. Briefly, the tissue homogenates suspended 10% by volume in Dulbecco's minimum essential medium (DMEM) and centrifuged. The supernatant was filtered with syringe filter (0.45 µm; ChmLab, Spain) and used to inoculate the MARC-145 cells grown in DMEM containing penicillin (100 units/ml), streptomycin (100 µg/ml) and amphotericin B (0.25 µg/ml) with 10% fetal bovine serum (FBS). After 2 h incubation at 37.5℃ and washing with phosphate buffered saline (PBS, pH 7.4), maintenance medium (DMEM with 5% FBS) was added. The suspicious virus candidates were in continuous passage with the same manner after 5~7 days of incubation until the cytopathic effect (CPE) was observed. When the CPE was observed, the PRRSV was identified by PRRSV-specific RT-nested PCR [20] and immunofluorescence assay (IFA). The IFA kit was kindly given from National Veterinary Research Quarantine Service (Korea).
RNA extraction from the PRRSV isolates and collected sera
In this study, the all collected sera were made to a pooled sample with 2 or 3 individual sera in the same age group. The pooled samples of 250 µl and culture medium of isolated virus (passage 3, both CP07-401-9 and CP07-626-2) was mixed with 750 µl of Trizol LS (Invitrogen, USA). In the case of virus infected tissues, 20% tissue suspension with PBS was made, and 250 µl of suspension was reacted with Trizol (Invitrogen, USA). After 15 min incubation, 200 µl of cold chloroform was added, followed by vortexing. The vortexed mixture was centrifuged at 12,000 g, 4℃ for 15 min and 450 µl of the supernatant was mixed with same volume of cold isopropyl alcohol. After precipitating overnight at -20℃, the solution was centrifuged at 12,000 g, 4℃ for 10 min and the final RNA pellet was acquired by washing with 75% ethanol and centrifugation. The RNA pellet was dissolved with 30 µl of 0.1% diethyl pyrocarbonate-treated distilled water.
cDNA synthesis and RT-PCR
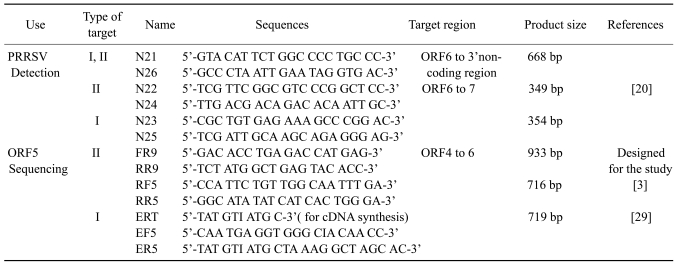
The primers used in this study were presented in Table 3. The cDNA was synthesized with a commercial M-MLV reverse transcriptase kit (Invitrogen, USA) following the manufacturer's instructions. The N26, RR9, ERT primers were used in the cDNA synthesis for virus detection and sequencing. To detect the viremic sera, a previously published RT-nested PCR method, which amplifies a part of ORF6 to ORF7 region, was used [20]. For the ORF5 sequencing of type II PRRSV, a modified RT-nested PCR was used based on the published RT-PCR method [3]. Briefly, FR9 and RR9 primers were newly designed based on the ORF4 and ORF6 regions to generate a 933 bp fragment to include the complete ORF5 sequence. These PCR products were further amplified with two published primers, RF5 and RR5, as a nested PCR which produces 716 bp fragment containing full ORF5 sequence.
The PCR reaction was performed with i-StarMaster mix PCR kit (iNtRON Biotechnology, Korea). The total mixture (20 µl) included 1 µl of each primer at a concentration of 10 pmol/µl, 2 µl of cDNA and 16 µl of master mix containing 250 mM dNTP, 2 mM Mg2+, K+ and NH4+ were added as salts along with 10 mM Tris-HCl (pH 9.0) to the PCR tube, which was coated with 2.5 Unit of i-star Taq DNA polymerase. The first PCR was performed at 95℃ for 5 min followed by 30 cycles of 95℃ 1 min, 55℃ 1 min, 72℃ 1 min, with a final extension at 72℃ or 10 min, and then held at 4℃.
The nested PCR was performed using 2 µl of first PCR products as a template along with FR5 and RR5 primers. The protocol was as follows: 95℃ for 5 min followed by 30 cycles of 95℃ 45 sec, 55℃ 45 sec, 72℃ 45 sec, with a final extension at 72℃ for 10 min, and then held at 4℃. For the ORF5 sequencing of type I PRRSV from the infected tissues, RT-PCR was performed as described in a previous paper [29]. The PCR products were electrophoresed in a 1% agarose gel containing ethidium bromide. The target bands were visualized using an ultraviolet lamp.
Sequencing and data analysis
For sequencing ORF5, a 716 bp fragment of type II PRRSV and a 719 bp fragment of type I PRRSV were purified using the QIAquick Gel Extraction kit (Qiagen Korea, Korea). Both strands of purified products were sequenced by Genotech (Korea). The full ORF5 sequences were analyzed using CLUSTALX v1.83 program and MegAlign software (DNAStar, USA) to determine the phylogenetic relationships and nucleotide similarity. The Neighbor-joining tree and was drawn using Kimura-two parameter as a distance estimation and percent frequencies of the groupings were determined after 1,000 bootstrap evaluation. The Maximum parsimony tree was also drawn as a consensus tree after 1,000 bootstrap evaluations.
Results
Pilot study about MLV vaccination and viremic status in 2007
The viremia of type II PRRSV was detected in both MLV-vaccinating and non-vaccinating farms (Table 4). In the non-vaccinating farms, most viremic pigs were found at 6 to 12 weeks of age. Even in the farms with MLV vaccination, PRRSV viremia was also detected regardless of vaccine schedule. The mean S/P ratio was relatively lower in the non-vaccinating farms than MLV-vaccinating farms at 9 and 12 weeks of age. However, mean S/P ratios were above 0.4 in all age groups, which meant that they were seroconverted for PRRSV.
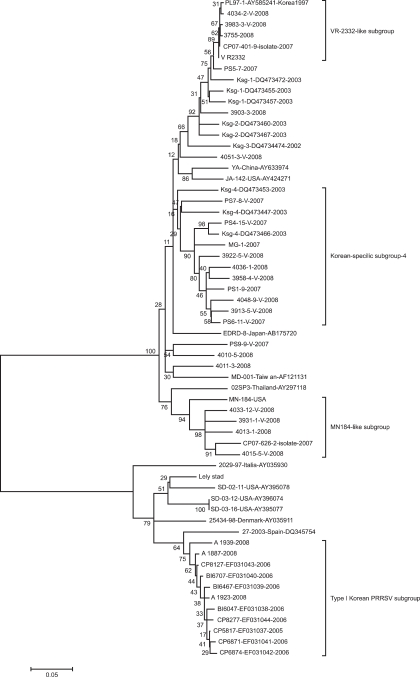
Phylogenetic relationship of total Korean PRRSV ORF5 sequences with those of other countries
The neighbor-joining tree was presented in Fig. 1. Korean specific subgroups (Ksg)-1, 2 and subgroup 3 were clustered near the VR-2332-like subgroup. The majority of the PRRSV samples collected from 2007 to 2008 were shown to belong to Ksg-4, which was previously reported by Cha et al. [7]. There was no subgroup-specific differences between viruses from MLV-vaccinating or non-vaccinating farms. The viruses of the Ksg-4 subgroup had 87.8-89.0% of nucleotide similarity with VR-2332 in the ORF5 region. The recent Korean PRRSVs from 2007 to 2008 made a novel subgroup which was not reported by Cha et al. [7] and were in the same cluster with MN184 strain from USA. The PRRSVs in this subgroup (MN184-like) had a similarity of 90.3~90.5% compared to MN184 and 84.9~87.2% compared to the MLV on nucleotides of the ORF5 region. The type I Korean PRRSVs were clustered with a 27-2003 strain from Spain with 89.1~91.7% of similarity. The Korean type I PRRSV had 84.9~88.4% of similarity in ORF5 when compared with a European-like PRRSV strain (SD-02-11, Genbank Accession no. AY395078) isolated from USA.
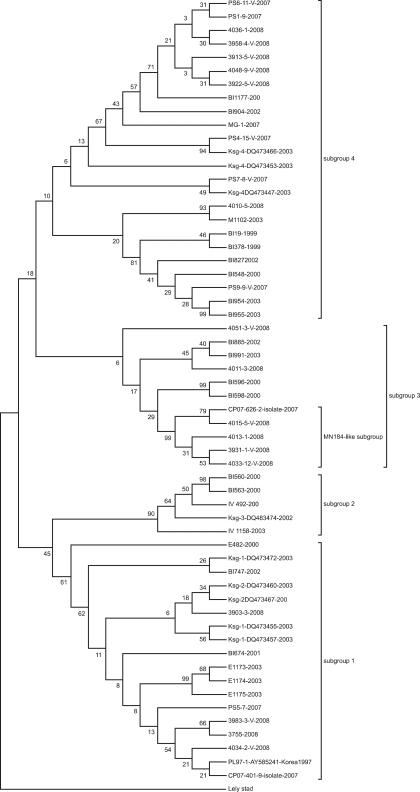
Evolution of type II Korean PRRSV from 1997 to 2008
To observe the evolutionary pattern of type II PRRSV from 1997 to 2008, a Maximum parsimony tree was drawn (Fig. 2). The type II Korean PRRSVs were shown to have been evolved into four subgroups, subgroups 1, 2, 3 and 4. Subgroup 1 included a PL97-1, a prototype of type II Korean PRRSV. The previously reported Ksg-1 and 2 subgroups were shown to belong to subgroup 1 and 24% (6/25) of recent type II Korean PRRSVs from 2007 to 2008, were also included in this group. Subgroup 2 containing Ksg-3 had only PRRSVs from samples taken as of 2000 to 2003. The subgroup 3 formed a novel cluster which was not reported before in Korea and had a MN184-like cluster. 28% (7/25) of recent PRRSVs belonged to this subgroup. Subgroup 4 consisted of 48% (12/25) of recent PRRSVs and was consistent with previously reports of the Ksg-4 subgroup.
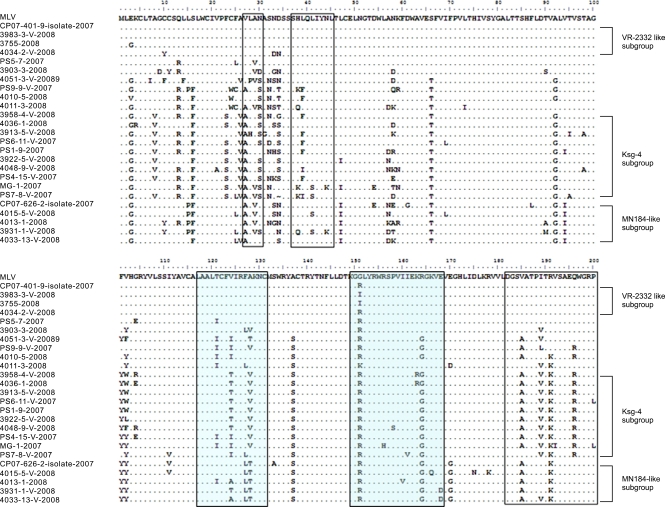
Analysis of deduced amino acid sequences of type II PRRSV ORF5 in the viremic sera
The amino acid sequences of 23 type II PRRSV ORF5 in the viremic sera taken from 2007 to 2008 were aligned to compare several epitope regions (Fig. 3). The Ksg-4-containing subgroup had at least one mutated sequences at H38, L39, L41 and N44 (K/Q38, F/I39, S41 and K44). In the case of the 3rd B-cell epitope (182~200), I189, R191 and Q196 were frequently replaced by V189, K191 and R196 in those groups. The MN184-like subgroup showed the replacement of V185 and R191 to A185 and K191. The first T-cell epitope (117~131) was also a variable region among Korean type II PRRSVs obtained from viremic sera. Most common was V124 and A128 being dominantly replaced by T/I124 and V/T128 in the Ksg-4-containing subgroup. Notably, the ORF5s in MN184-like subgroup had common mutations (FA127-128 → LT127-128) which were also observed in the isolate CP07-626-2 belonging to this subgroup. The second T-cell epitope (149~163) was more conserved than the first T-cell epitope (127~131) in the ORF5 sequences used in this study. The obvious differences of epitope regions were not observed between MLV-vaccinating or non-vaccinating farms, comparing the viruses in same subgroup.
Discussion
The viremia of heterologous virus after MLV vaccination had been reported in several papers. In previous studies, although MLV vaccination could contribute to improve the clinical outcome, sporadic viremia was as observed as ever after challenge with heterologous viruses [5,34]. Although clinical improvement in MLV-vaccinating farms was proved in those studies, those results could not reflect the actual situation in the field. Since the porcine circovirus type 2 (PCV2) became the one of the important pathogens causing the postweaning multisystemic wasting syndrome (PMWS, now one of the PCV-associated diseases), coinfection of other pathogens was the important factor for the disease appearance of PMWS. PRRSV was reported as the crucial factor of PMWS or severe wasting diseases when co-infected with PCV2 [1,12,33]. PCV2 was frequently detected in the wasting, diarrheic pigs and aborted fetuses in Korea [2,16,17]. Most farms in this study suffered from wasting during the nursery period (around 4-9 weeks of age) and several farms were shown to be infected with PCV2. Therefore, the viremia of vaccine-like virus or heterologous PRRSV observed even after MLV-vaccination might be the risk of wasting diseases when PCV2 was endemic in the farms, as viremia indicated the active replication of PRRSV [13].
In the pilot study, we could also observe sporadic viremia even in the seropositive herds of MLV-vaccinating farms applying vaccine schedules, such as sow-only and sow-piglet scheme. From this observation, this study focused on analyzing the PRRSVs obtained from the sera of viremic pigs in MLV-vaccinating or non-vaccinating farms. The PRRSV viremic pigs were frequently detected in both farms. Although high genetic variation was observed among the ORF5 of these viruses, there was no considerable differences between viruses from MLV-vaccinating or non-vaccinating farms. Since Korean swine farms were located not so far from the other farms and pigs were transferred on a national basis, frequent viral transmission 'from farm to farm' may be a factor for this observation. To understand vaccine pressure on PRRSV evolution, a long-term chronological study in the evolution of prevailing PRRSV after MLV vaccination should be performed on a farm to farm basis to reduce interfering factors such as transmission of foreign viruses.
There were some different phylogenetic patterns in this study which included the recent type II PRRSVs from 2007 to 2008. A new subgroup with 84.9~87.2% similarity of ORF5 compared to the MLV was identified. This subgroup consisted of only isolates collected from 2007 to 2008. In the neighbor-joining tree, they were shown to be closely related with MN184 strain from USA, showing 90.3~90.5% of similarity. Since USA is the major trade partner for the swine industry, these newly identified MN184-like viruses might have been introduced from USA one day and evolved in Korea independently.
The successful adaptation the CP07-626-2 strain, belonging to the MN184-like subgroup, on MARC-145 cells (passage 10), could be used to develop a new vaccine to control these newly emerging strains in Korea.
A Maximum parsimony tree of type II PRRSVs from 1997 to 2008 showed that the viruses had evolved after first introduction of VR-2332-like PRRSV (PL97-1 strain). At least 4 lineages were identified in the tree. Although the viruses in subgroup 2 were not detected recently, subgroups 1, 3 and 4 were still detected in Korea. Furthermore, the recent type II PRRSVs in subgroups 3 and 4 had high variations in their epitope regions of ORF5, including a novel B-cell epitope (3rd epitope) and two T-cell epitopes [11,38]. The neutralizing antibody fromation was known to be greatly affected by genetic variation and the amino acid sequences of ORF4 and ORF5 were more correlated with the neutralizing ability [19]. Therefore, these frequent substitutions in the epitope regions of ORF5 including neutralizing epitope also could be a cause of subsequent viremia, even in the MLV-vaccinating farms.
This study also confirmed that the type I PRRSV had been co-circulating with type II PRRSV in Korea. The type I PRRSV was primarily detected by RT-nested PCR-based typing which amplified the ORF6 to ORF7 region, and further confirmed by full sequencing of ORF5 in this study. The Korean type I PRRSV belonged to the one specific subgroup having 87.2~88.9% similarity in ORF5 compared to the Lelystad strain, the prototype. Notably, the ORF5 of type I Korean PRRSVs were similar with 27~003 strain of Spain, not the European-like PRRSV reported from the USA. This phylogenetic relationship suggested that the type I Korean PRRSVs were possibly introduced from Spain and had evolved into a single cluster. Notably, since the first report of type I PRRSV in Korea in 2006, the detection rate of type I PRRSV had been increasing in the field samples [18]. Because of the previously reported type-specific protection in the pigs [36,37], the use of type I PRRSV-based vaccine should be considered in Korea.
The viremia of heterologous PRRSV was frequently detected even in the MLV-vaccinating farms. These field viruses found in MLV-vaccinating farms were shown to have variable nucleotide substitutions on ORF5, similar to the non-vaccinating farms. Several substitutions of amino acids in the neutralizing epitope were also found in these viruses. The genetic diversity in type II PRRSV had been increasing from 1997 to 2008 in maximum parsimony tree. Furthermore, a newly emerging MN184-like subgroup appearing from 2007 was identified, and a circulation of type I Korean PRRSV was confirmed in this study. These complex situations observed from PRRSV molecular epidemiology in Korea reinforce the need for an additional vaccine strategy including a new vaccine development.

 XML Download
XML Download