

 PDF
PDF ePub
ePub Citation
Citation Print
Print


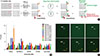
Breast cancer has been divided into five subtypes based on their intrinsic molecular entities: normal-like, luminal A, luminal B, human epidermal growth factor receptor 2-enriched, and basal-like [1]. Among them, basal-like breast cancer, commonly known as triple-negative breast cancer (TNBC), has attracted much interest due to its poor prognosis and lack of effective targeted therapies [2]. Intensive studies are, therefore, currently underway to identify the underlying molecular mechanism that drives the growth and metastasis of TNBC [3]. In addition, as early diagnosis enables early treatment and helps significantly delay disease progression and expand the overall lifetime of patients, the discovery of specific or selective biomarkers for early stage TNBC has become an attractive research area [4]. Nonetheless, there is still no specific biomarker that can provide a definitive diagnosis of TNBC. Current cancer screening is based on a combination of mammography and self-examination, but their prognostic value remains controversial [5]. No laboratory tests utilizing serum, saliva, or urine samples have been established to be effective in the primary diagnosis of breast cancer [5]. Thus, early stage biomarkers of TNBC to establish or develop precise early diagnostic methods that meet the criteria for an accurate, relatively noninvasive, reliable, and low-cost test remain to be discovered [67]. Recently, disease-specific autoantibodies derived from patient serum have become useful and cost-effective biomarkers, providing a potential gateway for diagnosing cancer [8]. Compared with other biomarkers such as DNA, RNA, or proteins, autoantibodies have several advantages in that they are structurally stable, specific, and present in large amounts in serum [7]. Thus, disease-specific autoantibodies are currently being investigated in a range of cancers to overcome the obstacles encountered for their early detection [8910]. In an effort to identify a useful early stage autoantibody biomarker in the sera from TNBC samples, we utilized a human protein microarray system containing 16,368 human recombinant proteins. Human serum proteome analysis demonstrated that approximately 3,700 separate proteins exist in a physiological state. One issue with using human serum to identify biomarkers is the high background protein content, which causes noise signals under certain experimental conditions [11]. To minimize background signals in the protein microarray, the amount of serum was adjusted so that up to 200 µg of serum proteins was used, which greatly reduced the interference by noise signals (data not shown). To identify autoantibodies in TNBC samples, we used the HuProt™ human proteome microarray v2.0 (CDI Laboratories Inc., Mayaguez, USA), each containing 16,368 unique full-length human protein antigens (Figure 1A). In total, 20 human serum samples (10 TNBC and 10 normal) were subjected to protein microarray analysis. This study was approved by the Institutional Review Board of Ajou University Hospital (IRB No. AJIRB-BMR-KSP-15-362) and written informed consent was obtained from all participants. The result of protein microarray was analyzed by GenePix analysis software (Molecular Devices, San Jose, USA) (Figure 1B). Briefly, microarray slides were blocked with filtrated dry milk and were then incubated with serum samples. After washing, the arrays were incubated with anti-human IgG (H+L) conjugated to AlexaFluor 546 (Cat. No. A-21445; Invitrogen, Carlsbad, USA). The arrays were then washed, dried, and immediately scanned with a GenePix 4000B Fluorescence Scanner (Molecular Devices). The GenePix analysis software determined that five autoantibodies, thioredoxin-like 2 (TXNL2), ADP-ribosylhydrolase like 1 (ADPRHL1), glycyl-TRNA synthetase (GARS), spinocerebellar ataxia type 3 protein (ATXN3) and solute carrier family 16 member 4 (SLC16A4), had a significantly higher expression in the TNBC samples than in the normal samples. The differential expression of these five autoantibody biomarkers is shown in Figure 1B. Among them, we selected the TXNL2 autoantibody biomarker not only because it showed the largest difference between the TNBC and normal samples but also because the TXNL2 autoantigen is known as a potential diagnostic indicator of breast cancer [12]. Figure 1C shows that the TXNL2 autoantibody specifically and strongly interacted with the TXNL2 antigen, suggesting that it is a highly specific biomarker for TNBC.
For independently verifying the TXNL2 autoantibody as a TNBC-specific biomarker, we attempted to purify the recombinant TXNL2 antigen from BL21-Gold(DE3)pLysS Competent Cells (Agilent, Santa Clara, USA). The purity of the TXNL2 protein is directly associated with the reliability of the TXNL2 autoantibody as a TNBC-specific biomarker. To obtain high-purity and sufficiently large yields of recombinant TXNL2, the full-length cDNA of TXNL2 was optimized for codon usage. Fortunately, we found that glutathione s-transferase (GST)-TXNL2 protein was successfully expressed and purified using a Glutathione Sepharose 4B (GE Healthcare, Little Chalfont, UK). Most GST-TXNL2 proteins were observed in the soluble fraction after sonication and migrated to ~63 kDa on an sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) as a single band (Figure 2A). Densitometric analysis by the ImageJ program (https://imagej.net/) indicated that GST-TXNL2 comprised 42% of the total proteins. As shown in Figure 2B, following affinity chromatography, the fractions containing GST-TXNL2 were further separated by using gel filtration with Superdex 75 prep grade resin (GE Healthcare) that displays a high resolution for proteins in the molecular weight range of 3,000 to 70,000 Da. Finally, we found that the final eluted fractions contained highly pure GST-TXNL2 (Figure 2C). The molecular weight and homogeneity of the sample were checked using a TSKgel-S3000SWXL (Tosoh Bioscience, Tokyo, Japan) (Figure 2D). Taken together, these data suggest that GST-TXNL2 protein might be a usable antigen for testing of the TXNL2 autoantibody as a TNBC-specific biomarker.
To validate the specificity of the TXNL2 autoantibody for TNBC, we employed a quantitative dot blot system. The dot blot method is the best technique for rapidly assessing the quantity of a target antigen across many samples at once. It is also a popular method for screening antibodies for target specificity. The feasibility of the dot blot analysis was verified by analyzing the GST protein and the specificity of a rabbit anti-GST antibody was evaluated (data not shown). For the dose-response curve, we serially diluted GST proteins from 0.1 µg to 2 µg under nondenaturing condition. Dot blot analysis yielded a linear curve between 0.2 µg and 1.8 µg of protein with a coefficient of determination (R2) of 0.999 when simple linear regression analysis was performed (data not shown). Beyond 2 µg, the signal intensity started to plateau. Based on the control experiment, we loaded 1.5 µg GST or GST-TXNL2 proteins onto the nitrocellulose membrane; we then incubated each membrane with normal or TNBC sera, which were used in the protein microarray experiment, as shown in Figure 2E. Dot blot data revealed that the amount of the TXNL2 autoantibody derived from each serum sample showed similar expression levels to those obtained using the protein microarray, indicating that the protein microarray data are reliable for detecting autoantibodies from sera (Figure 2F). In addition, we found that the TXNL2 autoantibody was more highly expressed in TNBC samples than in normal samples. Taken together, these results suggest that the TXNL2 autoantibody is a strong serum biomarker to detect TNBC.
Clinically, autoantibodies are currently being used to diagnose various immune diseases. However, they are also found in various cancers and neurological diseases, indicating that a diagnostic method using autoantibodies can be applied to a wide range of diseases beyond immunological diseases. That said, autoantibodies against cancer-related antigens may be detected in the sera of patients with various types of cancer, and there are still few ways to identify disease-specific autoantibodies from patient blood. Of these methods, protein microarrays have the advantage of being able to identify autoantibodies efficiently by using only a small amount of blood. For this reason, many research groups are now trying to identify autoantibodies as specific biomarkers for a number of diseases [13].
Recently, it has been reported that TXNL2 is also a TNBC-specific marker that is highly expressed in breast cancer patients [12]. Moreover, the same study determined that TXNL2 protein is directly linked to the growth and metastasis of breast cancer cells and is associated with decreased overall patient survival, suggesting that abnormal expression of TXNL2 is directly or indirectly linked to TNBC initiation or progression [12]. TXNL2 has a unique protein structure consisting of a Trx homology region in the N terminus, followed by two tandem repeats of the Grx domains, which play a role in reactive oxygen species (ROS) detoxification and cell viability [12]. Remarkably, the deletion of TXNL2 gene in mice leads to embryonic lethality [14], indicating its role in protecting cells against oxidative stress during embryogenesis. In addition, TXNL2 is a critical factor in regulating ROS levels and nuclear factor κB activity for controlling tumor development and metastasis in primary human breast cancers, demonstrating that it has essential roles in cancer cells [12]. In the current study, we identified the TXNL2 autoantibody as a TNBC-specific biomarker. Although the disease-specific linkage between autoantibodies and autoantigens is not clear, our data suggests that abnormal expression or dysfunctions of TXNL2 might be closely related to the generation of the TXNL2 autoantibody in TNBC.

 XML Download
XML Download